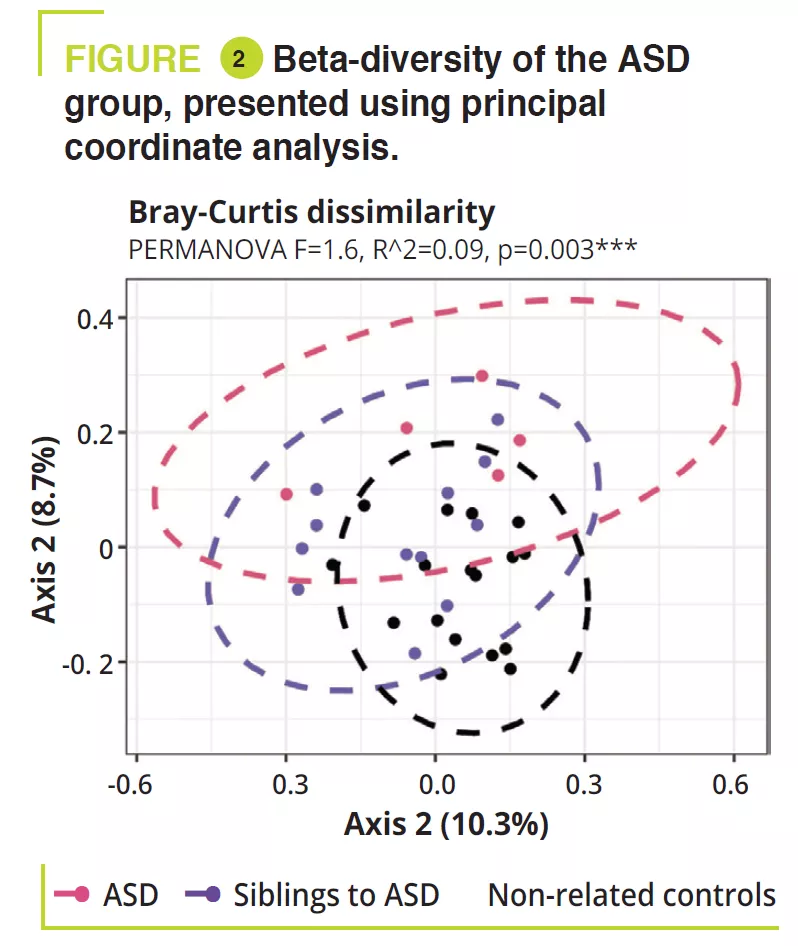
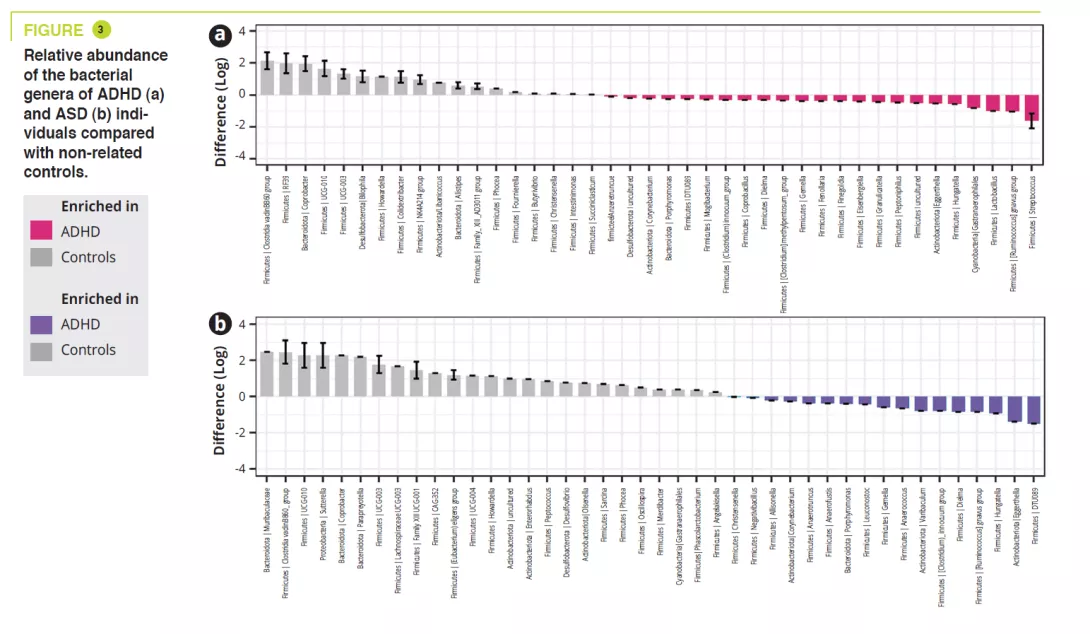
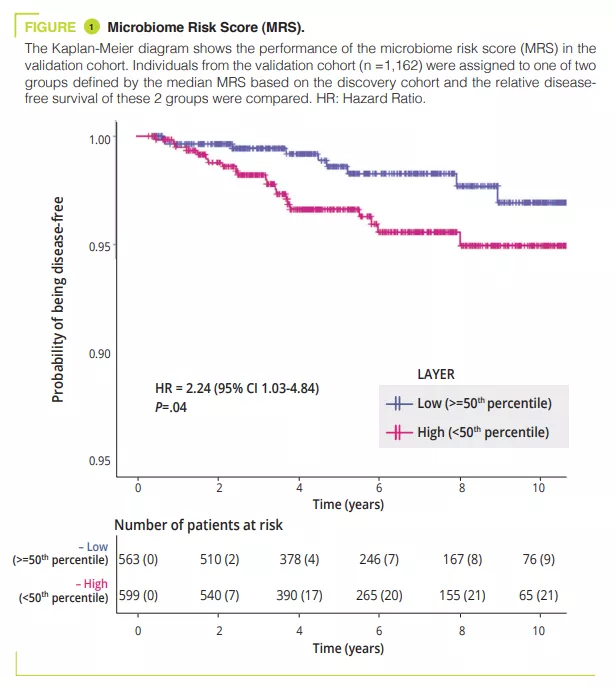
Role of gut microbiota in osteoporosis confirmed
A number of associations between the gut microbiota and bone density suggest the existence of a link between the gut flora and skeletal metabolism. Certain bacteria and metabolic pathways have been singled out.

If previous studies 1 are to be believed, there may be just one step from gut microbiota to skeletal health: certain gut microorganisms are thought to increase the production of T cells, which in turn stimulate the production of immune mediators and inflammatory cytokines, promoting osteoclastogenesis and bone loss in mice. Other studies point to mechanistic links involving the production of microbial short-chain fatty acids (SCFAs) and the metabolism of dietary components involved in bone metabolism (vitamins K, D, and complex polysaccharides). This research field even has a specific name: osteomicrobiology. The field has seen very few clinical studies, but an American team recently published one.
Two separate cohorts, one suspect in common
This research was based on the cohorts from two observational studies: 831 elderly men (mean age 84.2) from the Osteoporosis in Men Study (MrOS) 2 and 1,227 younger men and women (mean age 55.2) from the Framingham Heart Study (FHS) 3. An analysis of the data identified 37 microbial genera that appear to be involved in the FHS study (DTU089, Marvinbryantia, Blautia, and Akkermansia were negatively associated with bone density, while Turicibacterand Victivallis were positively associated) and 4 genera in the MrOS study (negative associations with Methanobrevibacter and DTU089 and positive associations with Lachnospiraceae NK4A136).
Thus, despite the difference between the two cohorts in terms of height, sex, and age, a common bacterium was associated with lower bone density in both cohorts: DTU089. DTU089 is known to be more abundant in those with low levels of physical activity and very limited protein intake, two factors unfavorable to bone health.

Meta-analysis
The researchers pooled the two cohorts to perform a meta-analysis. The results: a higher abundance of Akkermansia and DTU089 was associated with a less dense radius and tibia; conversely, a higher abundance of the Lachnospiraceae NK4A136 group and Faecalibacterium was associated with higher bone density.
The researchers also identified eight metabolic pathways associated with bone measurements, the most important of which involved the histidine, purine, and pyrimidine biosynthesis pathway. Previous experiments in mice had suggested a disturbance in purine metabolism in osteoporosis.
These results remain preliminary and require further studies to better understand the mechanisms by which certain bacteria can modify skeletal integrity, but they do support initial preclinical findings. Above all, they give greater hope that we may one day be able to modulate the gut microbiota and better protect bone health.
1. Okoro PC, Orwoll ES, Huttenhower C et al. A two-cohort study on the association between the gut microbiota and bone density, microarchitecture, and strength. Front Endocrinol (Lausanne). 2023 Sep 21;14:1237727. doi: 10.3389/fendo.2023.1237727.
2. https://mrosonline.ucsf.edu/
3. https://www.framinghamheartstudy.org/
 Gut microbiota and vitamin D: a promising team in the fight against osteoporosis?
Gut microbiota and vitamin D: a promising team in the fight against osteoporosis?
 Microgravity, microbiota, and bone density
Microgravity, microbiota, and bone density