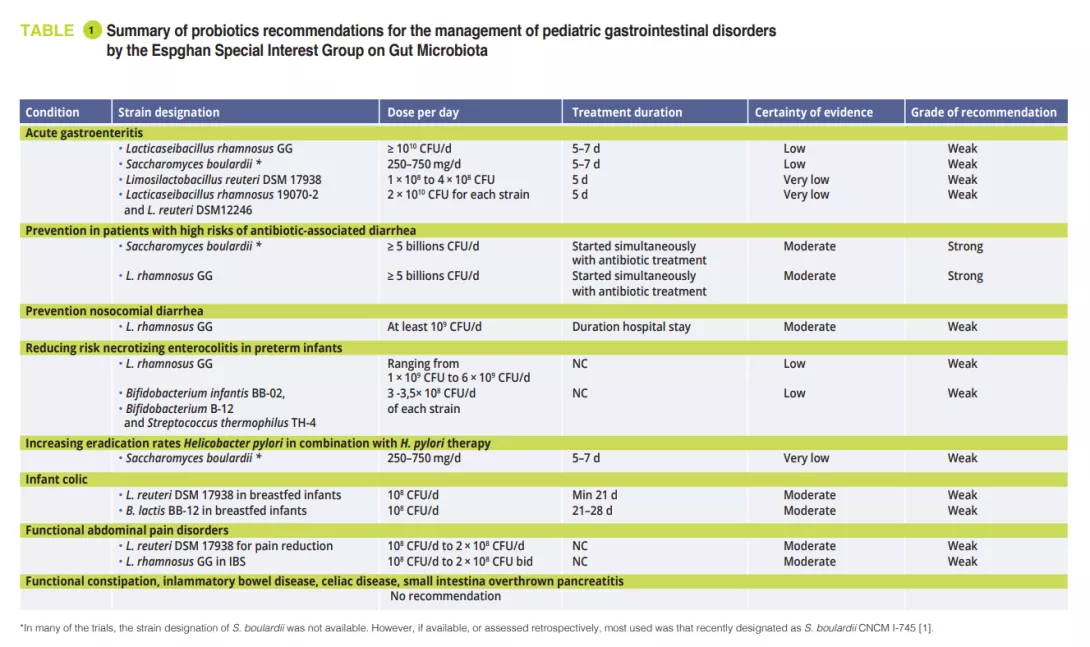
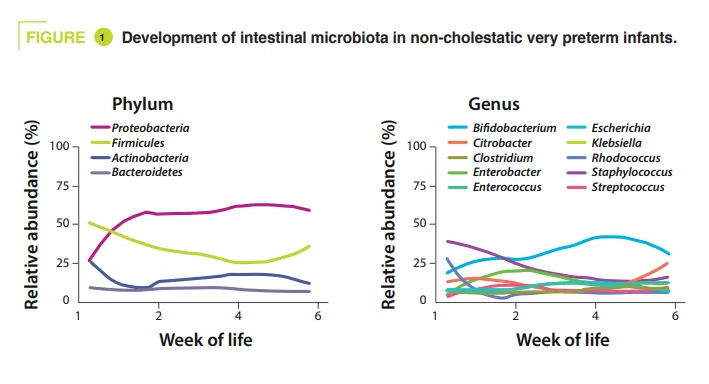
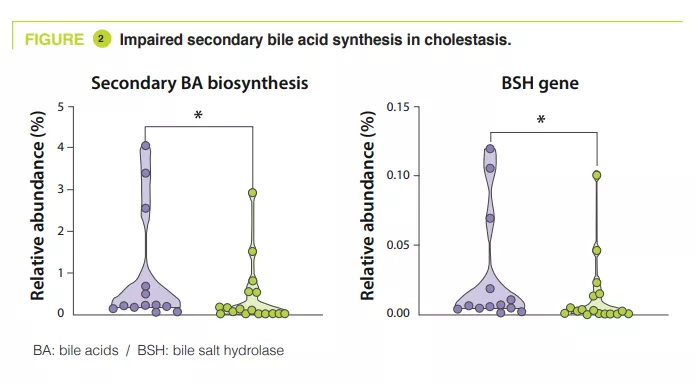
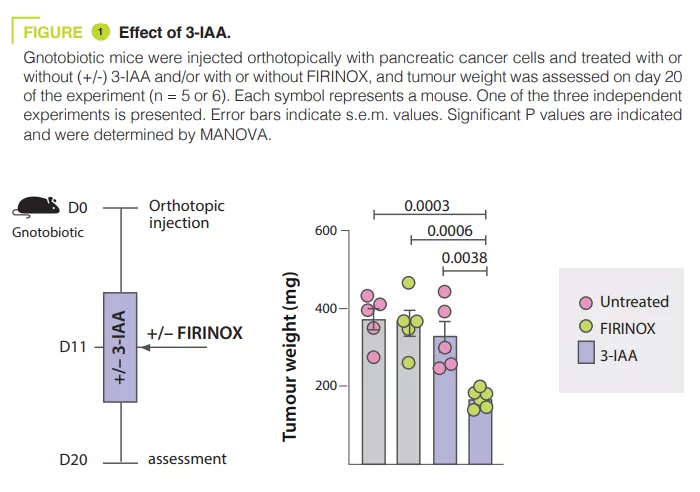
Gut Microbiota #19
By Pr. Satu Pekkala
Academy of Finland Research Fellow, Faculty of Sport and Health Sciences, University of Jyväskylä, Finland

Microbial metabolism of 5-ASA in inflammatory bowel disease
Inflammatory bowel disease (IBD) is treated with 5-aminosalicylic acid (5-ASA). However, more than half of the patients do not respond to the treatment. Previous studies have suggested that this is partly due to that 5-ASA can be metabolized by gut bacteria into N-acetyl 5-ASA that is clinically ineffective. In this elegant study Mehta et al. aimed at identifying the gut microbial enzymes that generate N-acetyl 5-ASA. The human stool samples were analyzed using multi-omics. The untargeted metabolomic analyses of the samples pre-5-ASA and post-5-ASA administration revealed potential microbial mediators of the anti-inflammatory effects of 5-ASA. These included a decrease in 2-aminoadipate, a bacterial metabolite that has been linked to greater oxidative stress. In addition, 5-ASA seemed to alter nicotinate metabolism, which may also explain some anti-inflammatory effects. The authors further sought to identify the microbial enzymes that potentially metabolize 5-ASA. By combining metatranscriptomics and metabolomics, they identified three Acetyl-CoA C-acetyltransferases (Acyl-CoA NAT) that associated with the N-acetyl 5-ASA levels in 5-ASA users. In addition, some thiolases were identified as potential candidate enzymes. The candidate enzymes were then heterologously expressed in Escherichia coli and their biochemical activity was measured. Firmicutes CAG:176 thiolase and Faecalibacterium prausnitzii acyl-CoA NAT were able to acetylate 5-ASA using acetyl-CoA. Ultimately, a metagenomic analysis of the stool samples revealed that the gut microbial 5-ASA-inactivating acetyltransferases associated with greater risk of treatment failure in 5-ASA users. Altogether, the findings of this study can help in advancing the possibility of microbiome-based personalized treatment of IBD.

Helicobacter pylori and colorectal cancer
Helicobacter pylori infection can lead to gastric cancer and increase the risk for colorectal cancer (CRC). However, mechanistic data on the latter is lacking. In this paper, Ralser and co-workers identify in a rodent model the underlying mechanisms of how H. pylori infection contributes to CRC. When the authors infected Apc mice that are excellent animal models bearing multiple intestinal neoplasia with H. pylori, an increased tumor burden in the small intestine and colon was observed. It is known that the T-cell immune response of the host contributes to gastric carcinogenesis, and therefore, the authors studied these responses in the intestines. They found a reduction in regulatory T cells and pro-inflammatory T cells as well as an increase in IL-17A, which is shown to be one of the main players in the immune response to H. pylori. The infected mice were characterized by higher abundance of so-called pro-inflammatory gut microbes, and mucus degrading bacteria, such as Akkermansia. By studying transcriptomic profiles of the intestinal epithelial cells, they found that H. pylori induced the activation of NF-κB and STAT3 pathways. Activation of these pathways has been previously shown also in CRC patients. Interestingly, germ-free mice infected with H. pylori scarcely showed activation of STAT3 signaling suggesting that H. pylori induced carcinogenesis in the small intestine is partly dependent on the gut microbiome. Ultimately, the authors show that H. pylori induced colorectal carcinogenesis can be prevented by eradication of the bacterium with antibiotics. The authors conclude that implementation of H. pylori status into preventive measures of CRC should be considered.
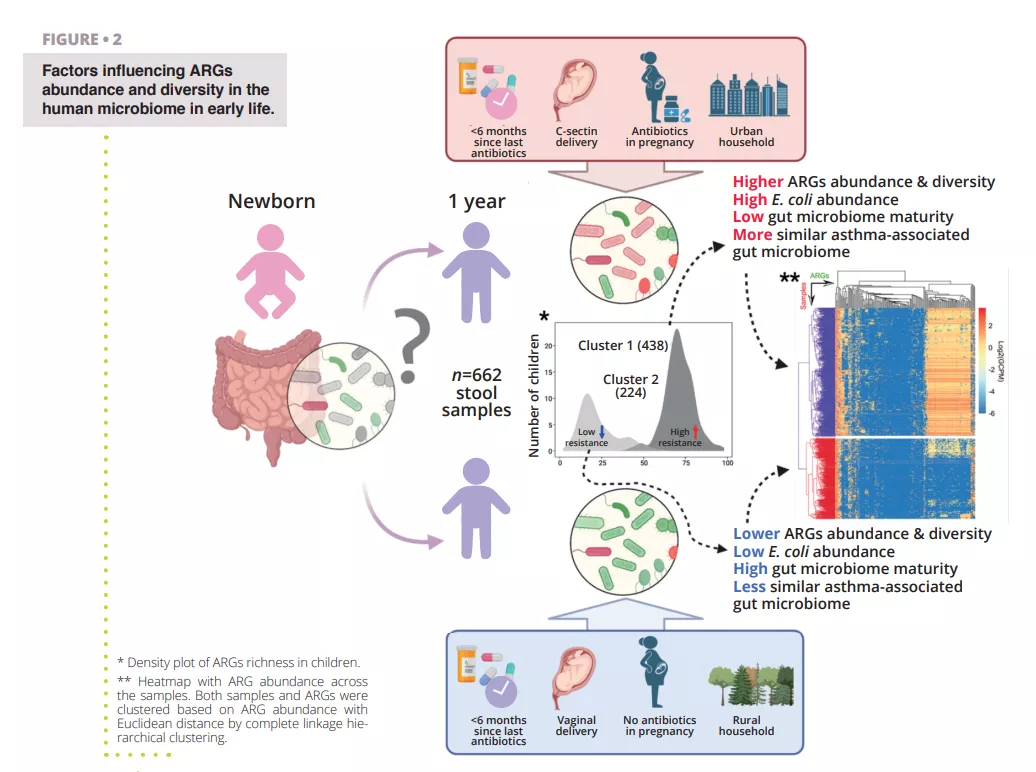
Viral diversity in the healthy infant gut
In infancy, the gut microbiome contributes to the maturation of the immune system to protect against chronic disease later in life. While it is known that bacteriophages (i.e., bacteria infecting viruses) can control the growth of bacterial populations, the gut viromes have not been extensively studied. By using metagenome sequencing, this study analyzed 647 viromes of a Danish cohort of 1-year-old infants. The first striking finding was that infant gut vOTUs were largely absent from gut virus databases. This suggests that the infant gut harbors specialized viruses distinct from the adult gut. The most predominant viral clades of the infants were largely undescribed. However, vertebrate-infecting ssDNA anelloviruses (Anelloviridae) and bacterial ssDNA microviruses (Petitvirales) were among the most abundant. In addition, virulent caudoviral families Skunaviridae, Salasmaviridae, β-crassviridae and Flandersviridae were also well represented in the infant viromes. Overall, temperate viruses were less prevalent than the virulent ones despite being found in more children. Family-level abundance was not significantly linked to phage lifestyle as determined by integrase as an indicator of a temperate lifestyle. However, temperate caudoviral families were genetically more diverse than the virulent families. Prediction analysis of the bacterial hosts of the viromes showed that Bacteroides, Faecalibacterium and Bifidobacterium were the three most prominent host genera in the infants’ gut. Of these, Bacteroides-infecting families were more often virulent and host specific. While no clear conclusions were provided, the study increases the knowledge on phage taxonomy and aids in the development of future infant gut virome research.