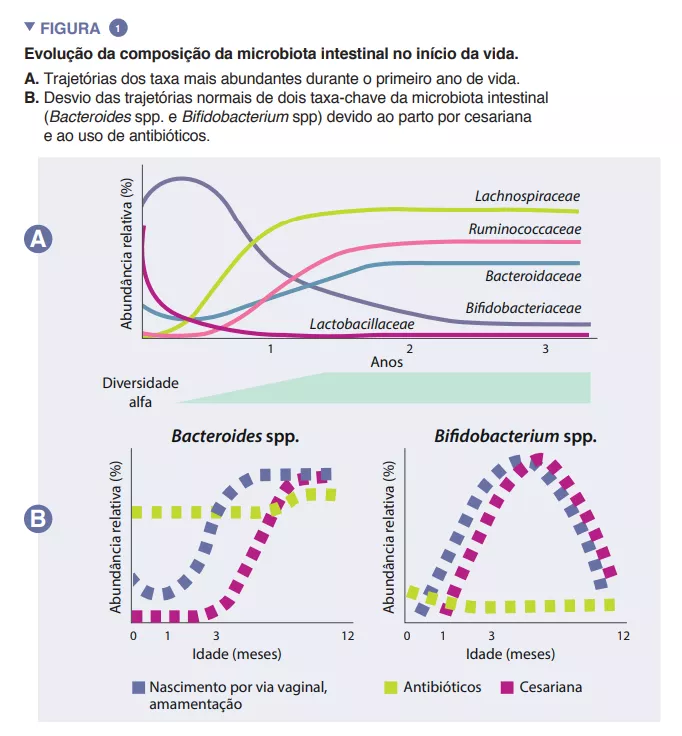
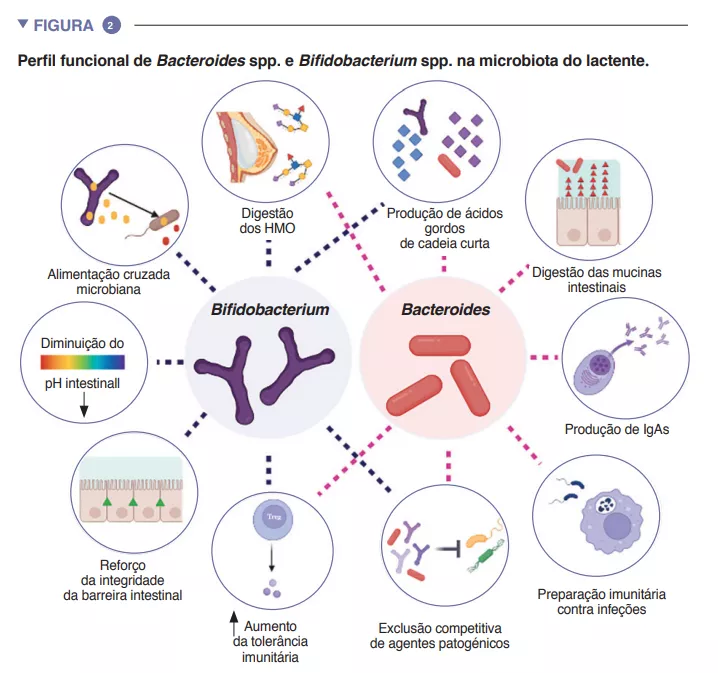
Da bactéria à crise de hiperfagia Quando a perda de F. prausnitzii conduz à crise de hiperfagia
Uma simples depleção de F. prauznitzii intestinal parece ser suficiente para criar hiperatividade em duas zonas do cérebro responsáveis por crises de hiperfagia. Como? Através da redução da produção de um metabolito e da estimulação do eixo intestino-cérebro pelo nervo vago. Um distúrbio que pode ser resolvido com a simples administração do probiótico ou do metabolito.
O consumo excessivo de alimentos gordos e açucarados é classicamente observado em pacientes que sofrem de distúrbios alimentares. Nas pessoas que sofrem de
(sidenote:
A hiperfagia bulímica
A hiperfagia bulímica é um distúrbio alimentar cujo diagnóstico se baseia em:
• critérios clínicos: comer em excesso pelo menos uma vez por semana, em média, durante 3 meses; sensação de falta de controlo sobre a alimentação.
• e na presença de 3 ou mais dos 5 critérios seguintes: comer muito mais rapidamente do que o normal; comer até ao ponto de se sentir desconfortável; comer grandes quantidades de alimentos sem sentir fisicamente fome; comer sozinho por vergonha; sentir-se enojado, deprimido ou culpado por ter comido demasiado.
O comportamento bulímico provoca um grande sofrimento.
Fonte: Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition Text Revision, DSM-5-TRTM, Feeding and eating disorders.
)
, estas crises são acompanhadas de infelicidade e excesso de peso, ou mesmo de obesidade. A microbiota intestinal parece estar implicada. Mas quais são os mecanismos envolvidos? Quais são os efeitos deste distúrbio alimentar na microbiota intestinal e desta última no comportamento? Para descobrir, os investigadores 1 decifraram passo a passo o eixo intestino-cérebro.
O distúrbio da hiperfagia bulímica afeta cerca de 3,5% das mulheres e 2% dos homens da população geral durante a sua vida. 2
Hiperfagia e disbiose intestinal
Tal como os humanos, após o stress e a dieta, os ratos têm mais tendência para comer biscoitos muito apetecíveis, aumentar o seu consumo de calorias e entregar-se à compulsão alimentar. Este comportamento parece estar ligado à microbiota intestinal dos roedores, cuja diversidade e riqueza são alteradas, com uma perda de Lactobacillus e Ruminococcaceae e um aumento de Bacteroides, Roseburia e Alistipes.

As experiências de transplante de microbiota fecal (FMT) sugerem que a flora dos ratos hiperfágicos pode estar empobrecida em bactérias protetoras: o FMT de ratos saudáveis faz com que os ratos hiperfágicos deixem de comer compulsivamente, sugerindo um regresso das bactérias protetoras. No sentido inverso, o FMT de ratos hiperfágicos para ratos saudáveis não induz o frenesim, sugerindo a ausência de bactérias que induzam este distúrbio.
Decifrar os mecanismos
Experiências complementares mostram que o frenesim alimentar nos ratos tem origem na desinibição do nervo vago que leva à hiperativação do eixo intestino-cérebro, passando pelo núcleo paraventricular do tálamo (associado à recompensa, à motivação e à homeostase energética) e pelo núcleo do trato solitário. O fator desencadeante poderá ser a baixa produção de um metabolito microbiano intestinal, o ácido quinurénico (KYNA), nos ratos hiperfágicos. A suplementação dos ratos com KYNA foi suficiente para os direcionar de volta para uma dieta equilibrada.
Tudo o que precisa de saber sobre o eixo microbiota-intestino-cérebro
E se bastasse um probiótico?
Para confirmar os resultados do modelo de murino, os investigadores analisaram amostras de fezes de 11 pacientes que sofrem de hiperfagia bulímica e 9 controlos saudáveis. A microbiota dos pacientes apresenta uma perda de Faecalibacterium prausnitzii (família de Ruminococcaceae) e uma diminuição dos níveis de KYNA. Por conseguinte, todas as razões levam a crer que uma redução de F. prausnitzii poderia acompanhar a redução do KYNA luminal e os distúrbios alimentares daí resultantes através do eixo intestino-cérebro. A hipótese parece ter sido confirmada em animais: a inoculação de F. prausnitzii em ratos hiperfágicos aumenta os níveis de KYNA, reduz a sua atração por biscoitos e a sua hiperfagia.
Por conseguinte, a toma de um suplemento de KYNA, o transplante fecal e/ou os probióticos poderiam desempenhar um papel no tratamento clínico de certos distúrbios alimentares, sob reserva de novas investigações, nomeadamente no ser humano. Entretanto, este estudo constitui um novo contributo para a compreensão do eixo intestino-cérebro.
2. Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition Text Revision, DSM-5-TRTM, Feeding and eating disorders.