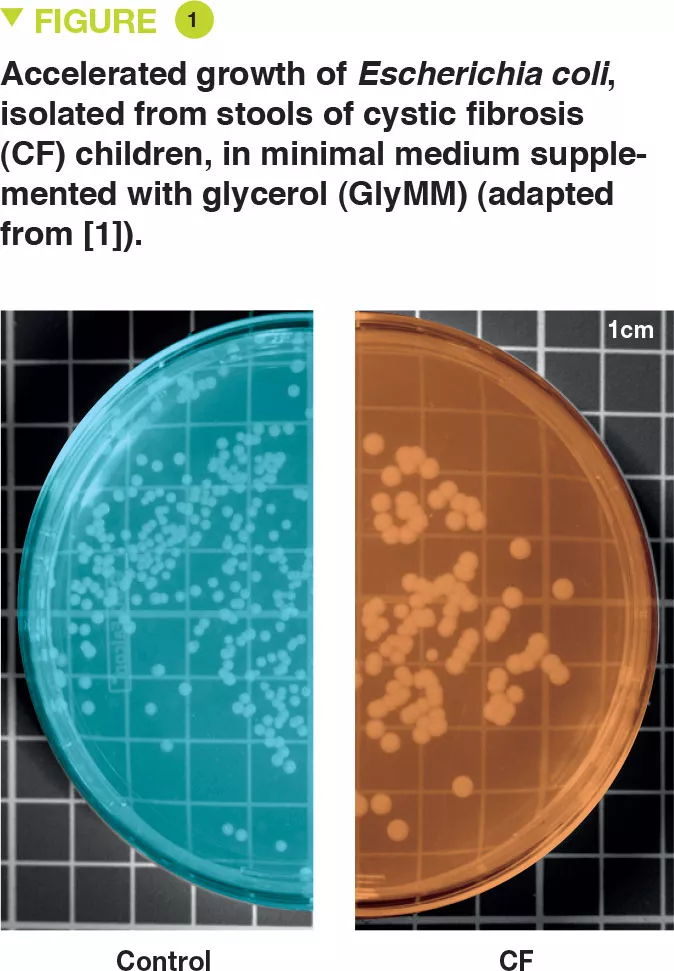
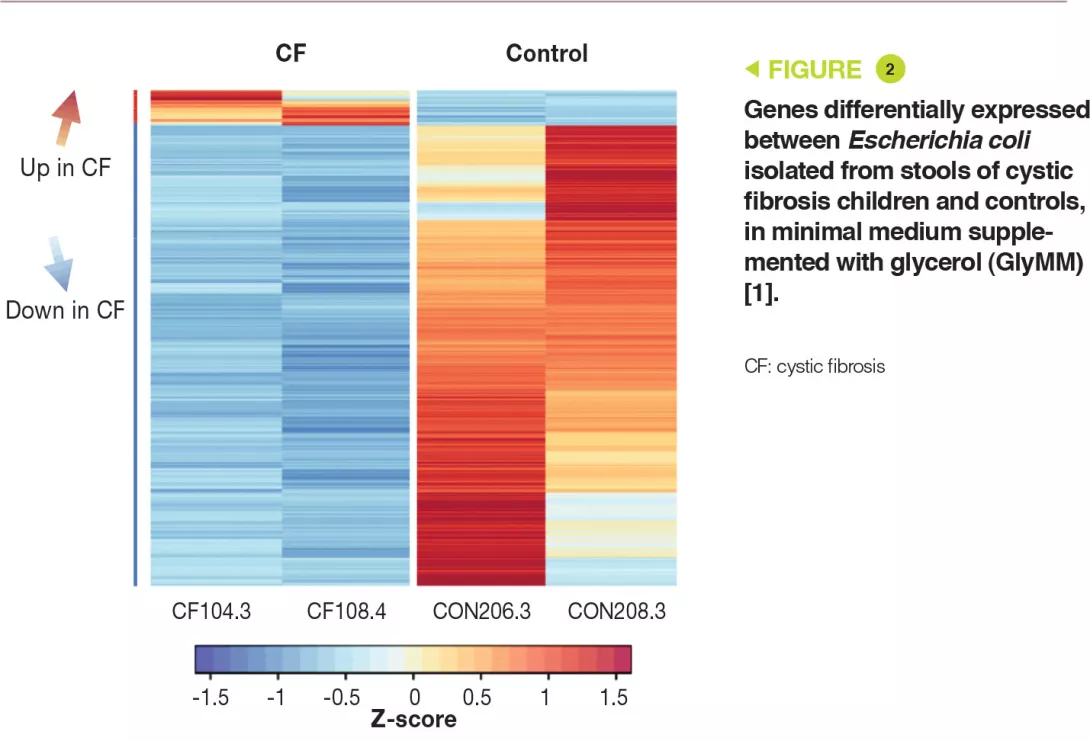
Gut microbiota composition and functions
Dr. Ralijic-Stojanovic stressed that sequence analysis of 16S rRNA has made it possible to confirm that the taxonomy of many cultured intestinal microorganisms was incorrect. For example, Clostridium difficile, which does not belong to the Clostridium butyricum genus, is in fact a distant relative of Clostridium perfringens, in contrast to what was previously assumed [1]. Dr. Ralijic-Stojanovic recalled that between the ages of 7 and 12 years, the GM is still different to that in adults. He concluded by stressing that GM composition is individual, specific, and stable, and may vary depending on age, diet, and lifestyle.
In his presentation, Dr. Bäckhed reported that, while the role of the GM in metabolism is well known (optimisation of caloric availability, intake of enzymes absent in humans, and the role in vitamin K synthesis and short-chain fatty acid production), some more recent publications have revealed that the level of butyrateproducing bacteria is reduced in patients with type 2 diabetes, increased levels of Prevotella improve glucose metabolism, and Christensenellaceae bacteria may be considered as an anti-obesogenic probiotic [2]. Dr. Bäckhed concluded that the GM should be considered as an environmental factor that contributes to host physiology and metabolism.
The GM is very complex, and despite the advances in recent years, all its secrets are yet to be revealed.
Gut microbiota and liver diseases
The role of the GM in liver diseases is being increasingly understood, and some authors have even suggested the existence of a “gut-liver axis”. Dr. Gasbarrini discussed the role of GM in liver inflammation and fibrosis, showing that severe alterations in GM have been observed in cirrhotic patients, with increased levels of Enterobacteriaceae, Veillonellaceae, and Streptococcaceae, and decreased levels of Clostridiacea, Lachnospiraceae, and Eubacteriaceae. Dr. Gasbarrini believes that an insufficient resilience, resulting in adaptation through the acquisition of a dysbiotic microbiota, may contribute to the onset of GM-associated chronic diseases. Intestinal barrier breakdown is the cornerstone to progression of fibrosis and severity of liver cirrhosis.
Another interesting aspect was also discussed: depending on the mechanism underlying liver injury, the GM may induce or prevent hepatic fibrosis. Possible ways to restore a healthy GM include GM modulation (diet, rifaximin, probiotics or prebiotics) or “reinitialisation” through faecal microbiota transplantation.
Dr. Kobyliak presented a poster [3] on a study conducted in patients with non-alcoholic fatty liver disease (NAFLD), who received a probiotic combined with flaxseed oil and wheat germs or a placebo for eight weeks. The results show that the concomitant administration of probiotics and omega 3 reduces liver fat and serum lipid levels, improves the metabolic profile, and reduces the chronic inflammatory state. Dr. Kobyliak concluded that modulating the GM through the use of probiotics is a new option in the management of NAFLD.
This confirms the influence of the GM on liver diseases and the possibility of alternative solutions through the use of probiotics.
Gut microbiota and chronic inflammatory bowel diseases (CIBD)
CIBDs are a heterogeneous group of immune-mediated chronic inflammatory diseases that affect the gastrointestinal tract. There are two main phenotypes of CIBD: ulcerative colitis (UC) and Crohn’s disease (CD). The relationship between the GM and CIBD is the subject of a growing number of publications.
Dr. Sokol discussed the pathogenesis of CIBDs and the fact that they are mediated by activation of the immune system through the GM in sensitive hosts under the influence of the environment. CIBD patients are known to have an abnormal microbiota with reduced diversity, which becomes increasingly reduced when the disease is active. Dr. Sokol stressed that there is an increase in Proteobacteria and a decrease in Firmicutes, which may or may not be correlated with the onset of disease. Thus, the level of adherent/invasive E. coli (Proteobacteria) is significantly increased in CD patients, but not in UC patients or healthy subjects. On the other hand, the level of Faecalibacterium prausnitzii (Firmicutes), which has antiinflammatory effects, is decreased in CIBD patients.
The environmental impact on GM is well known (mode of delivery, diet, antibiotics) and could also affect CIBD. Based on a report of a Danish cohort, Hviid et al. [4] observed a correlation between the number of antibiotic cycles received by a child and the risk of developing CIBD, which is greater for CD than UC.
Regarding pathogenesis, current controversy focuses on whether changes in the GM cause inflammation or vice versa; which came first: the chicken or the egg? Dr. Sokol believes that they are both at the same level, since the clinical manifestations of CIBD occur due to the implementation of a vicious circle between the GM and inflammation, and both may be the cause.
The data provided thus confirm the important role of GM bacteria in the pathogenesis of CIBD; however, our knowledge of the role of fungal GM in the pathogenesis of these diseases is limited. In this regard, Qiu et al. presented a poster [5] setting out the 15 main genera of fungi found in UC patients and healthy subjects (controls). For the Wickerhamomyces, Sterigmatomyces, and Penicillium genera, a positive correlation was observed with the expression of pro-inflammatory cytokines in the colonic mucosa, while the correlation was negative for Nigrospora. The authors concluded that the colonic fungal microbiota of UC patients is different to that of control subjects and that its alterations may be associated with mucosal inflammation and pathogenesis of UC.
Differences between the pathogenesis of CD and that of UC may be explained, in some cases, by the presence of a bacterial or fungal alteration of the GM.