Antibiotic-associated diarrhoea
The most frequent and best studied consequence of intestinal dysbiosis due to antibiotic intake is antibiotic-associated diarrhoea (AAD). AAD occurs during ±20% of all antibiotic courses and depends on the class of antibiotic, patient risk factors (host factors, hospitalization status, and nosocomial outbreaks), and the definition of AAD. AAD is defined as a change in stool frequency with at least three liquid stools/ day for two consecutive days, occurring during (early onset) or two to six weeks after antibiotic treatment (late onset), when no other cause can be identified (intercurrent viral or bacterial infection, laxative use, or other cause). The class of (broad-spectrum) antibiotics, duration of administration, and age of the patient are risk factors associated with the development of AAD. The administration of some probiotic strains, such as Lactobacillus rhamnosus and Saccharomyces boulardii, reduce the incidence and severity of AAD [4].
Antibiotics early in life
Antibiotics may have a much broader impact, especially when given perinatally or to young infants. Intrapartum antibiotics during Caesarean and vaginal delivery are associated with infant gut microbiota dysbiosis [5]. Dysbiosis acquired perinatally or during early life will induce long-term consequences. Maternal antibiotic treatment (during pregnancy and lactation) results in profound alterations in the composition of the microbiota in mothers and infants [6]. Prenatal antibiotics are associated with a larger body mass index (BMI) at the age of two years [7].
Antibiotics and weight
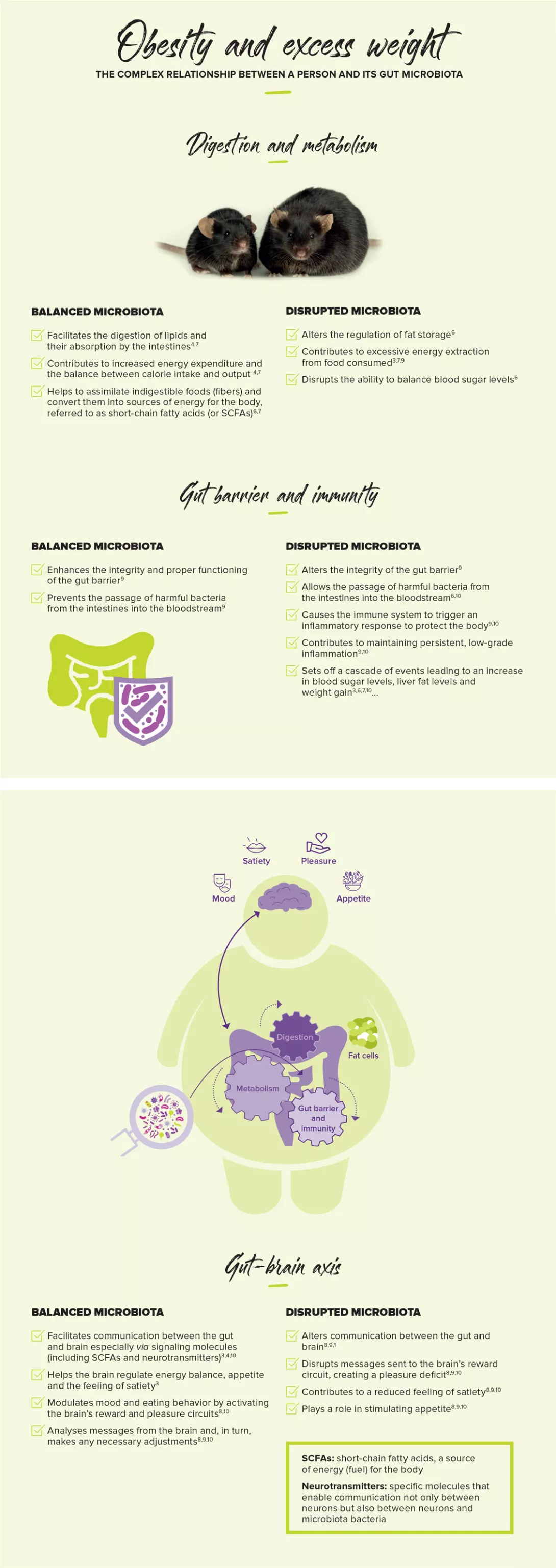
Sub-therapeutic doses of antibiotics have been used as growth promoters in animal farming since the 1950s [8]. The effect is more pronounced for broad-spectrum antibiotics, and is attenuated when animals are raised in sanitary conditions. Burgeoning empirical evidence suggests that antibiotics also affect human growth. As early as 1955, a randomized controlled trial in Navy recruits showed that a seven-week course of antibiotics led to significantly greater weight gain in the treated group compared with placebo [8].
There is a positive linear relationship between birth weight and BMI in six to seven-year-old children, which is present in both high and low-income countries [9]. The intestinal microbiota of infants is predictive of later BMI and may serve as an early indicator of obesity risk. Bifidobacteria and Streptococci, which are indicators of microbiota maturation in infants, are likely candidates for metabolic programming of infants, and their influence on BMI appears to depend on antibiotic use [10].
Antibiotic exposure before six months of age, or repeatedly during infancy, is associated with increased body mass in healthy children [11]. Repeated exposure to antibiotics early in life, especially β-lactam agents, is associated with increased weight and height [12]. Such effects may play a role in the worldwide childhood obesity epidemic and highlight the importance of judicious use of antibiotics during infancy, favouring narrow-spectrum antibiotics [11]. If causality of obesity can be established in future studies, this will further highlight the need for restrictive antibiotic use [12].
Administration of three or more courses of antibiotics before two years of age is associated with an increased risk of early childhood obesity [13]. In a cohort study, 6.4% children were obese at four years of age [13]. In this cohort, antibiotic exposure was associated with an increased risk of obesity at four years; the more antibiotic courses, the stronger the risk [13]. Children receiving antibiotics in the first year of life were more likely to be overweight later in childhood compared with those who were unexposed (32.4 versus 18.2% at 12 years of age; p=0.002) [14]. Repeated exposure to broad-spectrum antibiotics at ages 0 to 23 months is associated with early childhood obesity [15].
However, some studies have reported contradictory results. Exposure to antibiotics within the first six months of life compared with no exposure was not associated with a statistically significant difference in weight gain up to seven years of age [16].
Gut microbiome - immunity & food allergy
Symbiotic host and microbe interactions are critical for host metabolic and immune development. Early microbiota colonization may influence the occurrence of metabolic and immune diseases [1].
A clear association was found between early-life antibiotic use (three or more courses) and milk allergy, non-milk food allergy, and other allergies in a longitudinal data analysis of 30,060 children [17]. The associations became stronger for younger age and differed according to antibiotic class [17].
Maternal use of antibiotics before and during pregnancy was shown to be associated with an increased risk of allergy to cow’s milk in the offspring, and persisted after adjusting for putative confounders [17]. In children, the risk of allergy to cow’s milk increased with increasing number antibiotics used from birth to diagnosis (test for trend; p<0.001) [18]. Maternal intrapartum antibiotic prophylaxis has been shown to have a significant impact on the infant faecal microbial population, particularly in breastfed infants [19]. Intrapartum antibiotic administration was reported to result in a significant reduction in Bifidobacterium spp. strains [20]. The reduced abundance of these beneficial microorganisms, together with the increased amount of potentially pathogenic bacteria, may suggest these infants are more exposed to gastrointestinal or general health disorders later in life [20].
Antibiotics and the respiratory tract
Antibiotics given during the first week of life is a risk factor for allergic rhinitis and wheezing, while early introduction of solid foods, such as fish, and living on a farm are protective factors for the development of later allergic disease. Antibiotics taken by the infant during the first year of life is associated with an increased risk of asthma [21]. The strength of the association differs with the class of antibiotics, correlating with their effect on the gastrointestinal microbiome [21].
Antibiotic exposure has been associated with increased risk of asthma at three and six years of age [22], in the presence or absence of a lower respiratory tract infection during the first year of life [22]. The adverse effect of antibiotics was particularly strong in children with no family history of asthma (p [interaction] =0.03) [22]. Antibiotic intake was also a risk factor for a positive allergy blood or skin test. According to a systematic review published in 2011, exposure to antibiotics in the first year of life is a significant risk factor to develop asthma. Retrospective studies provided the highest pooled risk estimate for asthma compared with database and prospective studies. Respiratory infections, later asthma onset (asthma at or after two years), and exposure to antibiotics during pregnancy are all independent risk factors.
Antibiotic use in the first year of life is associated with the development of transient wheezing and persistent asthma [23]. A dose-response effect was observed; with five or more antibiotic courses, the risk to develop asthma increased significantly (p<001). There is no association between antibiotic use and late-onset asthma [23]. Antibiotic use in the first year of life is associated with an increased risk of early-onset childhood asthma, starting before three years of age. Reverse causality and protopathic bias may be confounders of this relationship [23].
Antibiotics and IBD
Exposure to antibiotics throughout childhood is associated with IBD, and this relationship decreases with increasing age of exposure to antibiotics. Exposure before one year of age was shown to have the highest risk, decreasing at five and 15 years, although antibiotics at the age of 15 still represented a significant risk factor to develop IBD [24]. Each antibiotic course increased the IBD hazard by 6% (4%-8%) [24]. As with any observational study, causality cannot be inferred and the possibility of data confounded by indication, due to prescription of antibiotics to children with intestinal symptoms of yet undiagnosed CD, should also be considered [25]. Antibiotic use is common in childhood and its potential as an environmental risk factor for IBD warrants scrutiny [25]. Antibiotic exposure has been reported to be significantly associated with Crohn’s disease, particularly in children, but not significantly associated with ulcerative colitis [26].
Antibiotics and diabetes
Exposure to a single antibiotic prescription was not shown to be associated with a higher adjusted risk of diabetes [27], whereas treatment with two to five antibiotic courses was associated with an increase in risk of diabetes for penicillin, cephalosporins, macrolides, and quinolones. The risk increased with the number of antibiotic courses. No association between exposure to anti-virals or anti-fungals and risk of diabetes was demonstrated [27]. Exposure to antibiotics is likely to increase the risk of type 2 diabetes [28]. However, these findings may also represent an increased demand for antibiotics due to an increased risk of infections in patients with yet-undiagnosed diabetes [28]. Antibiotic exposure in childhood is generally not associated with a risk of developing type 1 diabetes [29]. Future studies should investigate the effects of multiple exposures to broad-spectrum antibiotics during the second year of life.
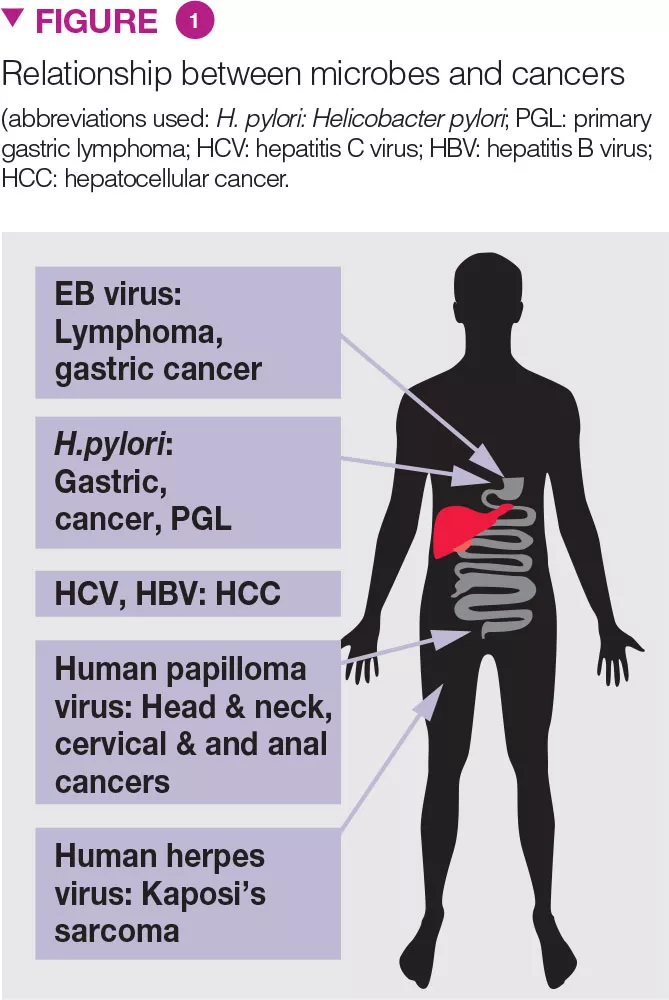
Antibiotics and malignancies
For gastro-intestinal malignancies, the use of penicillin has been shown to be associated with an elevated risk of oesophageal, gastric, and pancreatic cancers [30]. The association increased with the number of antibiotic courses. The risk of lung cancer increased with the use of penicillin, cephalosporins, or macrolides. The risk of prostate cancer increased modestly with the use of penicillin, quinolones, sulphonamides, and tetracyclines. The risk of breast cancer was modestly associated with exposure to sulphonamides. There was no association between the use of anti-virals or anti-fungals and risk of cancer [30].