Le microbiote vaginal
Comment prendre soin de son microbiote vaginal ?
Le vagin abrite des centaines de bactéries1. Penchons-nous sur leur fonctionnement et voyons pourquoi il est important de prendre soin de son microbiote.

21% Seule 1 femme sur 5 affirme connaître le sens exact du terme « microbiote vaginal »
Qu’est-ce que le microbiote vaginal exactement ?
Le microbiote vaginal (ou flore vaginale) est composé de centaines de bactéries et d’une quantité plus faible de champignons (Candida) qui vivent dans le vagin1.
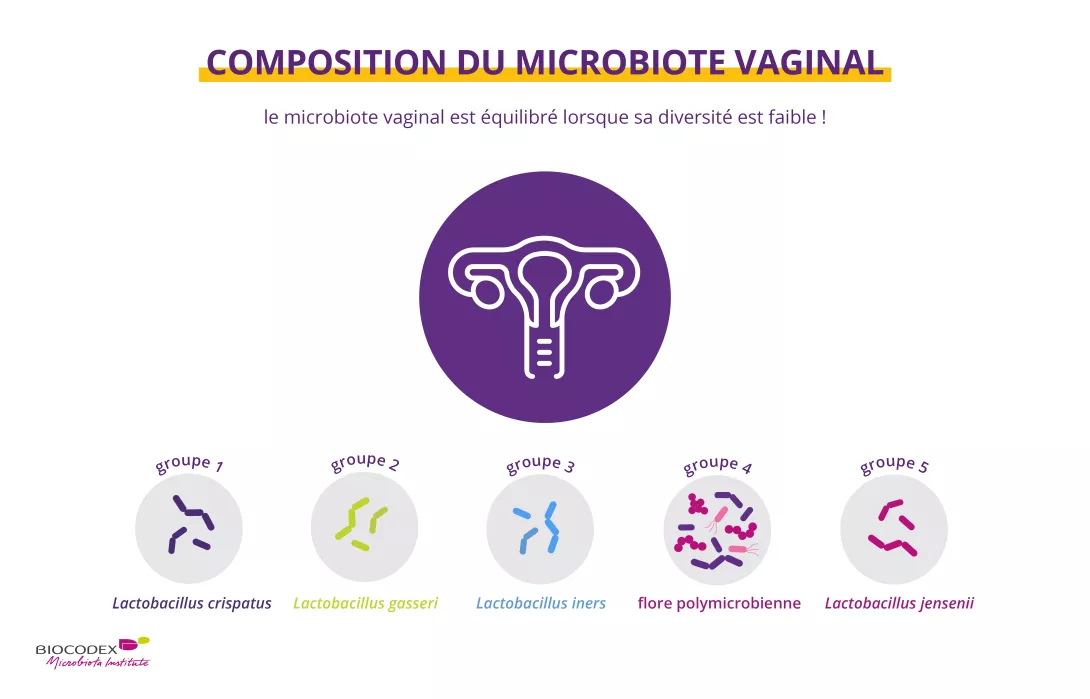
Chez la plupart des femmes, contrairement au microbiote intestinal, le microbiote vaginal est équilibré lorsqu’il montre une faible diversité (environ 200 espèces de bactéries) et lorsque les lactobacilles (bactéries en forme de bâtonnets) sont prédominants1.
Toutes les femmes ont un microbiote vaginal, mais aucune n’a le même. Pour le moment, cinq sortes principales de communautés bactériennes ont été décrites1,2:
-
quatre communautés dominées par les (sidenote: Lactobacilles Bactérie en forme de batônnet, dont la caractéristique principale est de produire de l’acide lactique. C’est pour cela que l’on parle de « bactéries lactiques ». Ces bactéries sont présentes chez l’homme au niveau des microbiotes oral, vaginal, intestinal, mais aussi sur les plantes ou chez les animaux. On peut les consommer dans les produits fermentés : produits laitiers comme certains fromages et yaourts, mais aussi des d’autres types d’aliments fermentés : les cornichons, la choucroute etc.. Les lactobacillus sont aussi consommés dans les probiotiques, certaines espèces étant reconnues pour leurs propriétés bénéfiques. W. H. Holzapfel et B. J. Wood, The Genera of Lactic Acid Bacteria, 2, Springer-Verlag, 1st ed. 1995 (2012), 411 p. « The genus Lactobacillus par W. P. Hammes, R. F. Vogel Tannock GW. A special fondness for lactobacilli. Appl Environ Microbiol. 2004 Jun;70(6):3189-94. Smith TJ, Rigassio-Radler D, Denmark R, et al. Effect of Lactobacillus rhamnosus LGG® and Bifidobacterium animalis ssp. lactis BB-12® on health-related quality of life in college students affected by upper respiratory infections. Br J Nutr. 2013 Jun;109(11):1999-2007. ) (Lactobacillus crispatus, L. gasseri, L. iners, ou L. jensenii) ;
-
une communauté caractérisée par une faible teneur en lactobacilles, voire par leur absence totale.
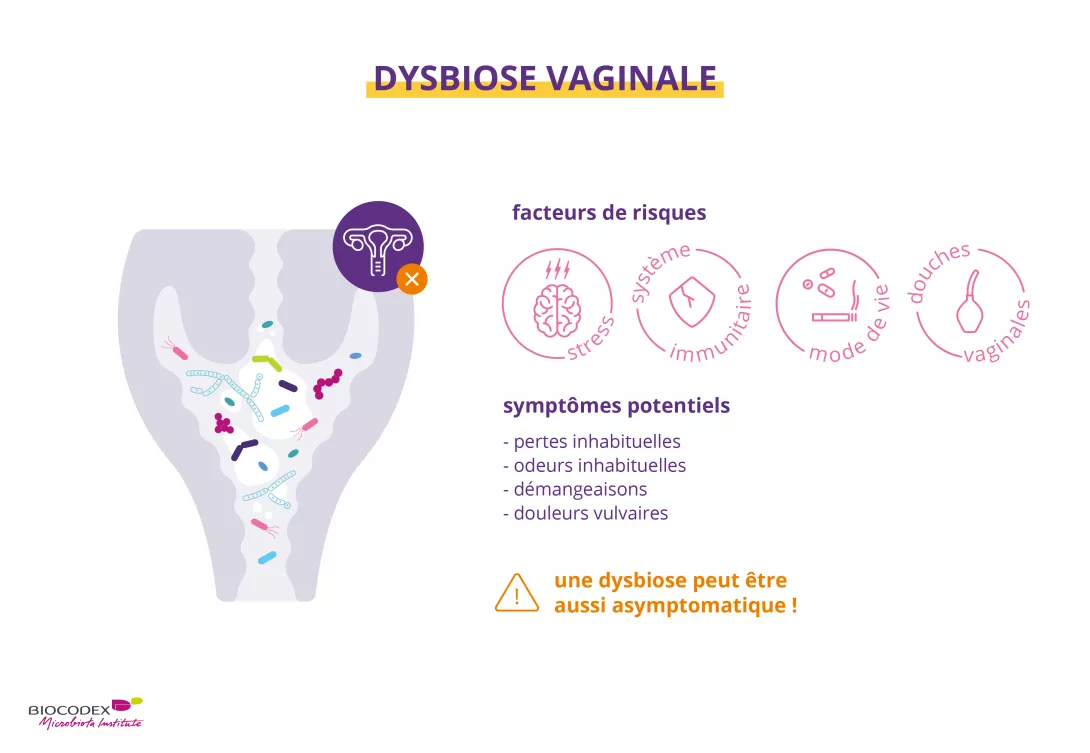
Le microbiote vaginal est une communauté dynamique soumise à l’influence de différents facteurs tels que l’origine ethnique, les hormones sexuelles, la contraception hormonale, le comportement sexuel, l’utilisation de douches vaginales, le régime alimentaire, le tabagisme, l’environnement social (par exemple, le cadre de vie) et les gènes1,3.
Cependant, la flore vaginale n’est pas totalement isolée de son environnement. L’anus et l’entrée du vagin sont très proches l’un de l’autre, si bien que des bactéries intestinales peuvent coloniser le vagin. L’intestin constitue donc un réservoir naturel de lactobacilles pour le vagin, ce qui est important pour l’équilibre de la flore vaginale5,6,7.
Comment le microbiote vaginal évolue-t-il tout au long de la vie ?
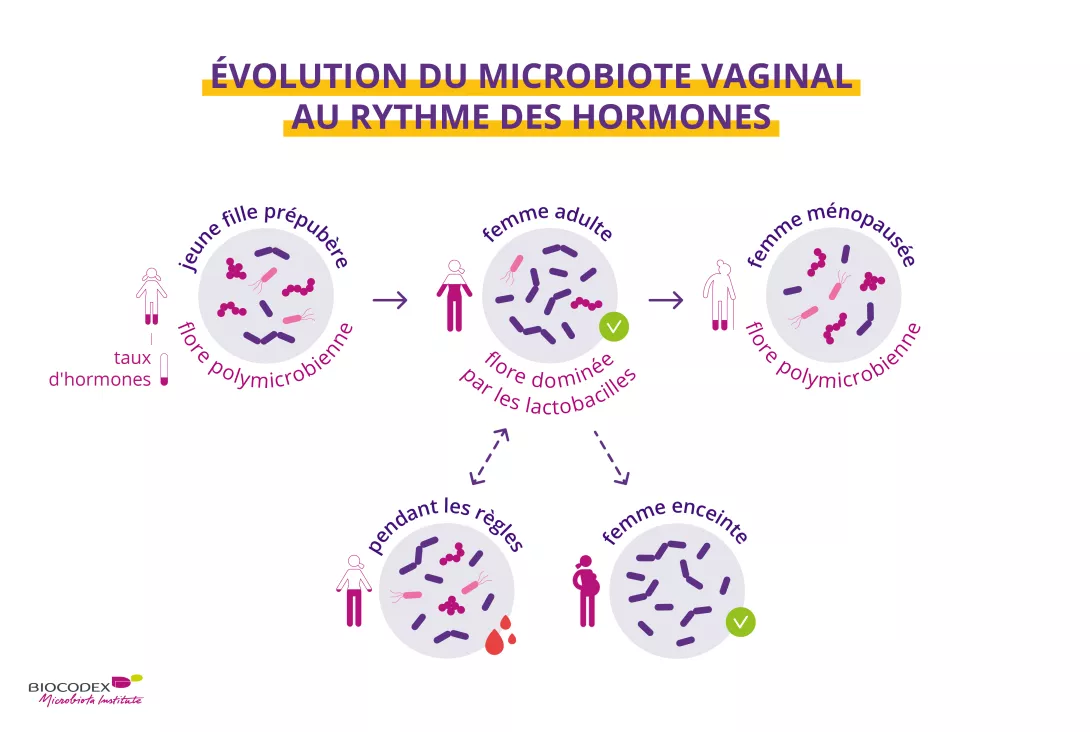
Le corps évolue tout au long de la vie et le microbiote vaginal ne fait pas figure d’exception. La composition du microbiote vaginal subit d’importants bouleversements de l’enfance à la ménopause en passant par l’âge adulte1. Des changements hormonaux marquent le rythme de notre vie et ont également un impact sur le microbiote vaginal. Par exemple, la menstruation modifie temporairement la diversité du microbiote vaginal8. Celui-ci joue également un rôle important lors de l’accouchement1,10. Au cours de la grossesse, des changements physiologiques permettent l’adaptation du corps de la mère au fœtus et réciproquement9. Chez la femme enceinte, le microbiote vaginal est plus stable, moins riche et moins diversifié9, tandis que les taux élevés d’œstrogènes assurent la prépondérance absolue des lactobacilles1,8. Enfin, à la ménopause, le microbiote vaginal trouve un nouvel équilibre10.
Pourquoi le microbiote vaginal joue-t-il un rôle important pour la santé ?
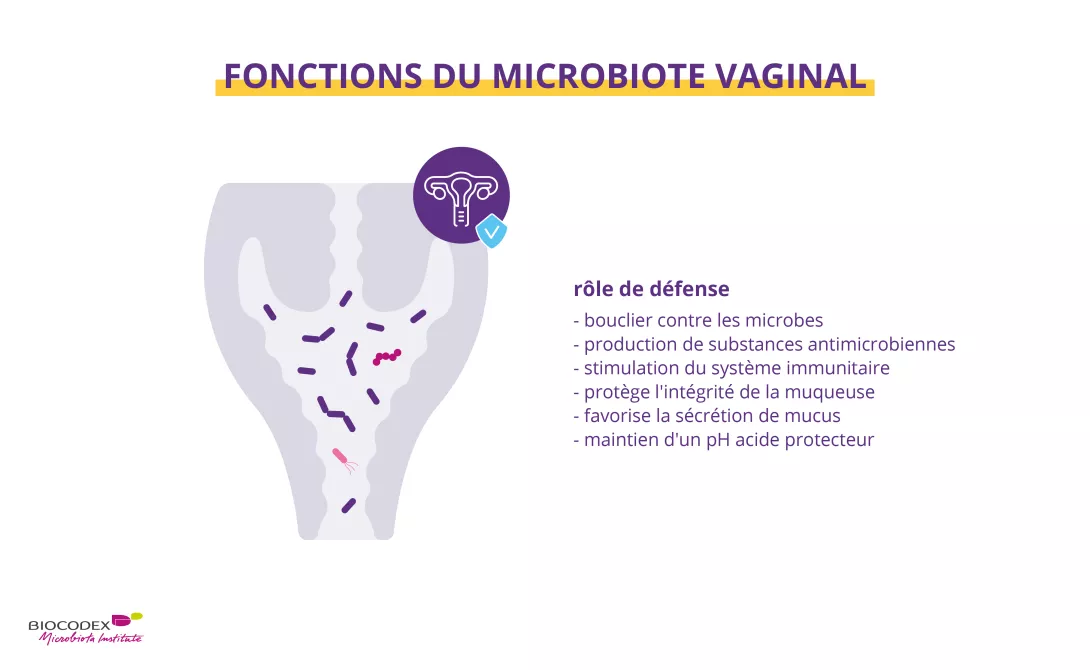
Les bactéries du microbiote vaginal contribuent à maintenir un environnement vaginal sain1. Certaines de ces bactéries, notamment les lactobacilles, empêchent les
(sidenote:
Microorganismes
Organismes vivants qui sont trop petits pour être vus à l'oeil nu. Ils incluent les bactéries, les virus, les champignons, les archées, les protozoaires, etc… et sont communément appelés "microbes".
Source : What is microbiology? Microbiology Society.
)
pathogènes de s’installer dans le vagin. Pour expliquer ce phénomène, plusieurs mécanismes ont été proposés :
- en produisant de l’acide lactique, le microbiote favorise un environnement acide (pH ≤ 4,5) qui est hostile pour de nombreux pathogènes11,12
- certaines molécules produites par le microbiote, comme le peroxyde d’hydrogène (H2O2) ou des substances antibactériennes (bactériocines), attaquent les bactéries, les virus et les champignons considérés comme des intrus11,12 le microbiote agit comme une barrière, empêchant les pathogènes de s’installer sur les parois du vagin. La présence de lactobacilles accélère la rénovation de l’épithélium auquel les pathogènes pourraient essayer d’adhérer11,12
- le microbiote facilite la production par l’épithélium vaginal d’un mucus protecteur qui maintient les pathogènes à distance11,12
- en stimulant le système immunitaire de la femme, le microbiote améliore sa capacité à repousser les attaques de pathogènes11,12
30% 3 femmes sur 10 savent que le microbiote vaginal est équilibré lorsque sa diversité bactérienne est faible.
Quelles sont les maladies associées à un déséquilibre du microbiote vaginal ?
Stress, maladie, excès d’hygiène (douches vaginales), médicaments (antibiotiques, etc.), alcool, tabac… tous ces facteurs peuvent affecter la composition du microbiote vaginal.8, Lorsque celui-ci est déséquilibré, il se produit ce que l’on appelle une « (sidenote: Dysbiose La « dysbiose » n’est pas un phénomène homogène : elle varie en fonction de l’état de santé de chaque individu. Elle est généralement définie comme une altération de la composition et du fonctionnement du microbiote, provoquée par un ensemble de facteurs environnementaux et liés à l’individu, qui perturbent l’écosystème microbien. Levy M, Kolodziejczyk AA, Thaiss CA, et al. Dysbiosis and the immune system. Nat Rev Immunol. 2017;17(4):219-232. ) » 8,11.
La dysbiose vaginale se caractérise par la perte de prédominance des lactobacilles – les bactéries les plus importantes de la flore vaginale –, ce qui ouvre la voie à des microorganismes opportunistes capables de coloniser le vagin8,11. Si leur présence est souvent accompagnée de pertes vaginales, de prurit et d’une sensation de brûlure, ou encore d’une odeur de poisson, elle peut tout aussi bien être asymptomatique8.
La dysbiose vaginale peut être associée à:
- une vaginose bactérienne liée à la colonisation par des bactéries pathogènes1
- une candidose provoquée par la prolifération d’un champignon8
- une diminution de la fécondité11
- une augmentation du risque d’accouchement prematuré1
A savoir
Il convient de garder à l’esprit que les femmes souffrant de vaginose bactérienne sont plus exposées au risque d’infections sexuellement transmissibles (IST) comme l’herpès, le papillomavirus, le SIDA (VIH) ou encore d’infections bactériennes (gonorrhée, chlamydiose, trichomonase)3,14.
Compte tenu des facteurs ayant un impact direct sur notre microbiote vaginal, comment pouvons-nous en prendre soin?
Il est essentiel de prendre soin de son microbiote vaginal. L’hygiène intime quotidienne est primordiale pour éviter les dysbioses. Comme les réseaux sociaux véhiculent pléthore de fausses informations, il est important de bien comprendre les choses qu’il faut faire et celles qu’il ne faut pas faire.

Au quotidien :
Si les douches vaginales ne sont pas recommandées car elles altèrent la flore vaginale, le lavage extérieur de la vulve avec un gel conçu spécialement pour l’hygiène intime15 contribue à réduire l’accumulation indésirable de pertes vaginales, de sueur, d’urine et de contaminants fécaux16.
Afin de contribuer à la bonne santé du microbiote vaginal...
... l’hygiène reste bien sûr nécessaire15 mais elle n’est pas suffisante. Un certain nombre d’options existent déjà pour le microbiote vaginal et d’autres en sont au stade d'essai :
- Probiotiques: les probiotiques sont des microorganismes vivants qui, lorsqu’ils sont administrés en quantité appropriée, produisent des effets bénéfiques sur la santé de l’hôte17,18. Ils sont notamment capables de réduire ou de corriger sans danger les déséquilibres du microbiote. Les probiotiques féminins administrés par voie orale ou vaginale peuvent contribuer à restaurer la flore vaginale, à améliorer les symptômes et à réduire le risque de récidive de différentes infections vaginales13,19,20,21. Cela est vrai aussi bien pour les femmes en âge de procréer que pour les femmes ménopausées13,20,21.
- Prébiotiques: les prébiotiques sont des fibres alimentaires non digestibles qui exercent des effets positifs sur la santé et qui sont utilisés de manière sélective par les microorganismes bénéfiques du microbiote de l’hôte22,23. Les produits spécifiques associant probiotiques et prébiotiques sont appelés « symbiotiques »24. Les prébiotiques féminins sont censés favoriser la prolifération des lactobacilles et contribuer à la normalisation de l’acidité vaginale19,25,26.
Toutes les informations contenues dans cet article proviennent de sources scientifiques autorisées. Ne perdez pas de vue toutefois que ces informations ne sont pas exhaustives. Consultez ici l’ensemble des études d’où nous avons tiré ces informations.
L'Observatoire International des Microbiotes
12 Kovachev S. Defence factors of vaginal lactobacilli. Crit Rev Microbiol. 2018 Feb;44(1):31-39.
15 Bohbot JM, Rica E. Microbiote vaginal, la révolution rose. Editions Marabout. 288 p.
BMI-21.12
 L’impact des contraceptifs sur nos microbiotes ? C’est quitte ou double !
L’impact des contraceptifs sur nos microbiotes ? C’est quitte ou double !
 Vaginose bactérienne : bientôt une greffe de microbiote vaginal ?
Vaginose bactérienne : bientôt une greffe de microbiote vaginal ?