



Magazine
Profilaxis antibiótica durante el parto frente a infección por egb
Revisión de prensa
Por el Pr. Ener Cagri DINLEYICI
Servicio de Pediatría, Facultad de Medicina, Universidad Eskisehir Osmangazi, Eskisehir, Turquía
Summary
Off

Migrated content
Désactivé
Updated content
Désactivé
Foco sobre la semana asia pacífico de enfermedades digestivas
De vuelta del congreso
Por el Pr. Uday C Ghoshal
Departamento de Gastroenterología, SGPGI, Lucknow, India

La comprensión del sistema gastrointestinal (GI) y de sus trastornos ha mejorado recientemente gracias a los conocimientos sobre la microbiota intestinal (MI) y la disbiosis [1]. Durante este congreso se presentaron varios aspectos de la MI, que mencionamos en este artículo.
Introducción a la microbiota intestinal
Considerada como un auténtico órgano, la microbiota intestinal posee 10 veces más células (1014) que las células humanas de nuestro organismo (1013) [2]. Las funciones de la MI incluyen la digestión de los alimentos, el metabolismo de medicamentos y toxinas, la síntesis de vitaminas, un efecto barrera frente a las bacterias patógenas, una modulación de las funciones del sistema nervioso central, endocrino e inmunológico [2]. Si consideramos las diversas funciones de la MI, podemos asumir que su alteración está relacionada con varias enfermedades, y que su modulación puede ser beneficiosa.
La función de la microbiota intestinal en el cáncer colorrectal
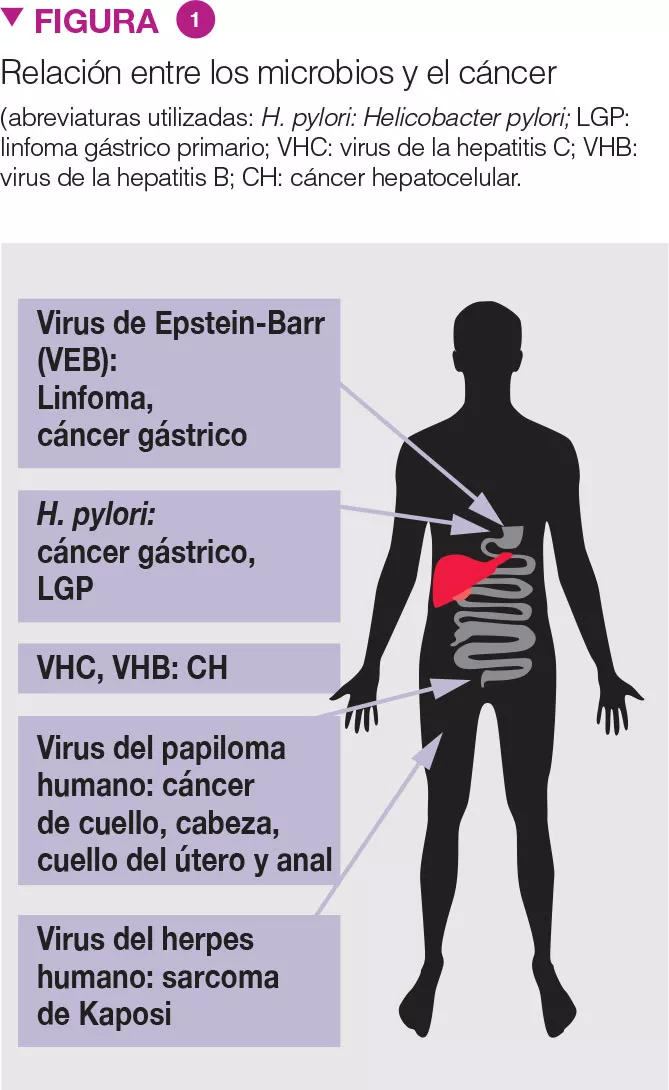
La asociación entre microorganismos y cáncer es conocida (Fig. 1) [3]. Recientemente, nuevos datos sugieren el rol de la disbiosis en el CCR. La microbiota fecal de pacientes con pólipos de colon se parece a la de pacientes con CCR. Mientras que Clostridium spp., Bacteroides spp. y Bifidobacterium spp. están asociados al CCR, las bacterias productoras de ácido láctico (p. ej., Lactobacillus spp. y Eubacterium aerofaciens) están negativamente asociadas con el CCR. La producción de metano y de H2S (sulfuro de hidrógeno) asociada a la MI, y la presencia de Streptococcus bovis, podrían desempeñar una función en el desarrollo del CCR. Además, se ha sugerido que la obesidad podría estar relacionada con la MI, y esta constituir un factor predisponente de CCR.
Imagen
La microbiota intestinal y la obesidad
La extracción de calorías de los alimentos no solo depende de la función digestiva del intestino delgado, sino también de la extracción de calorías malabsorbidas por la microbiota del colon. Mientras que la presencia de Firmicutes está asociada con una mayor extracción de calorías, las Bacteroidetes tienen el efecto contrario [4].
Se ha evidenciado una microbiota fecal diferente entre personas obesas y personas no obesas. En un estudio retrospectivo de cohortes en el Reino Unido, de 21 714 lactantes, 1306 (6 %) eran obesos a la edad de 4 años. En un análisis de regresión logística y tomando en cuenta el contexto familiar, la exposición a antibióticos antes de los 2 años de edad se asoció al desarrollo de la obesidad, y el número de ciclos de antibióticos se correlacionaba con el desarrollo de la obesidad [5].
Otros síndromes metabólicos
La esteatosis hepática no alcohólica (EHNA) y el síndrome metabólico pueden estar asociados a una disbiosis, incluyendo un aumento cuantitativo de las bacterias del intestino superior [6]. Un estudio no controlado y tres estudios de casos y controles demostraron que el crecimiento excesivo bacteriano en el intestino delgado estaba asociado con la EHNA [6]. Dos estudios demostraron una abundancia relativa menor de Bacteriodetes, y una abundancia mayor de C. coccoides y de Prevotella, en pacientes con EHNA. Una mayor extracción de calorías de carbohidratos complejos no absorbidos, una resistencia a la insulina, y una producción endógena de alcohol puede contribuir a la patogenia de la EHNA debido a la disbiosis.
La MI desempeña un papel importante en el metabolismo de la glucosa, la resistencia a la insulina, la diabetes, y tiene una incidencia en su tratamiento. Los pacientes con diabetes tienen microbiotas fecales distintas de las de la población control [4]. Se demostró que la MI es un factor importante de regulación de los niveles de glucosa después de la ingesta de diferentes alimentos, independientemente del ejercicio físico y/o del estilo de vida [7]. Es posible que la metformina, un antiglicémico oral, funcione parcialmente mediante la alteración de la MI. A pesar de que los estudios sobre la función de la MI en la enfermedad coronaria son escasos y poseen resultados diversos, están surgiendo nuevos datos que sugieren su implicación en esta enfermedad.
Uso indebido de antibióticos en asia
El uso de antibióticos en Asia es elevado, y la implementación de políticas para el uso debido deja que desear, lo que implica un riesgo de aparición de superbacterias resistentes a los antibióticos. Este uso indebido se explica por una disponibilidad sin restricciones, y un uso en indicaciones inadecuadas como, por ejemplo, resfriados o gastroenteritis aguda. El uso de probióticos, cuando corresponde, puede ayudar a contener el uso indebido de antibióticos.
Manipulación de la microbiota intestinal con agentes diferentes de los antibióticos
Aunque la modulación de la MI por parte de la rifaximina ya se conoce, los probióticos y el trasplante fecal son potencialmente interesantes para tratar los trastornos asociados a la disbiosis. Los probióticos pueden ser prescritos de forma concomitante en caso de tratamiento de eliminación de Helicobacter pylori. Hay metaanálisis que muestran que la administración concomitante de probióticos puede restaurar la eubiosis, y aumenta la tasa de erradicación, debido a menores efectos adversos y a un mejor cumplimiento [9]
A partir de 23 ensayos controlados aleatorizados, el Colegio Estadounidense de Gastroenterología recomendó la utilización de probióticos para mejorar el meteorismo y la flatulencia en el síndrome del intestino irritable[8]. Un análisis de Cochrane demostró la utilidad de los probióticos en la prevención de la diarrea asociada a Clostridium difficile [9]. Un metaanálisis demostró que los probióticos de diferentes especies provocan y mantienen la remisión de la rectocolitis hemorrágica, aunque los datos de la enfermedad de Crohn son limitados [10].
Orientaciones futuras
Con el objetivo de formar un consorcio de la región Asia Pacífico sobre la MI, similar a los grupos europeos y norteamericanos, y de analizar los datos actuales en la región Asia Pacífico, se ha desarrollado y publicado un consenso [9]. Las principales conclusiones del consenso son las siguientes: aumenta la evidencia que respalda el potencial terapéutico de los probióticos para modular las funciones gastrointestinales, y aliviar los síntomas de estos trastornos, pero se necesita más investigación [9].
Bibliografia
Summary
Off

Migrated content
Désactivé
Updated content
Désactivé
Informe sobre el 30a congreso del grupo europeo de helicobacter
De vuelta del congreso
Por el Pr. Francis Mégraud
Laboratorio de Bacteriología Hospital Universitario Pellegrin, Burdeos, Francia

Summary
Off

Migrated content
Désactivé
Updated content
Désactivé
El contenido de heces de lactantes con cólicos induce hipersensibilidad visceral en ratones
Artículo comentado - niño
Por el Pr. Emmanuel Mas
Gastroenterología y nutrición, Hospital de Niños, Toulouse, Francia

El comentario del artículo original de Eutamène et al. (Neurogastroenterol Motil 2017)
Poco se comprende de la fisiopatología de los cólicos infantiles, a pesar de que diversos estudios informan sobre la disbiosis intestinal en lactantes con cólicos. Nuestro objetivo fue evaluar la hipótesis de que la disbiosis asociada a los cólicos está relacionada con una hipersensibilidad visceral desencadenada por una alteración del contenido intraluminal.
Se estudiaron muestras fecales de siete lactantes con cólicos y de otros siete sin cólicos. Se infundieron sobrenadantes fecales (SF) en cólones de ratones C57/Bl6 (n=10/muestra). Posteriormente se evaluó la sensibilidad visceral en los animales mediante el registro de la respuesta muscular abdominal a la distensión colorrectal (DCR) a través de una electromiografía (EMG). Se evaluó la actividad de las proteasas de cisteína y serina en los SF con sustratos específicos. Se analizó la composición de la microbiota fecal de lactantes mediante la extracción de ADN y de la pirosecuenciación del gen 16S rARN.
Los SF de los lactantes con cólicos desencadenaron una actividad mayor en la EMG que los SF de los lactantes sin cólicos, en respuesta a los mayores volúmenes de DCR y en general, ya que se evaluó el área bajo la curva de la EMG en todos los volúmenes de la DCR. El tiempo de llanto de los lactantes está fuertemente relacionado con la actividad de la EMG de los ratones. La variedad de microbiota y la diversidad filogénica aumentaron en el grupo con cólicos, sin mostrar alteraciones destacadas en la composición microbiana. En el grupo con cólicos solo aumentaron Bacteroides vulgatus y Bilophila wadsworthia. La abundancia de Bacteroides vulgatus está positivamente relacionada con la sensibilidad visceral. No se encontraron diferencias en las actividades de la proteasa.[1]
¿Qué es lo que ya se sabe sobre este tema?
Los cólicos del lactante forman parte de las alteraciones funcionales gastrointestinales y se definen según los criterios de Roma. Su fisiopatología todavía es poco conocida, si bien se sospecha de un mecanismo intestinal doloroso. Hay estudios que sugieren que una perturbación de la microbiota intestinal, un aumento de la permeabilidad intestinal, y una inflamación intestinal de bajo grado, estarían implicados en la hipersensibilidad visceral. Esos factores intervienen en la fisiopatología del síndrome del intestino irritable. En este síndrome, una anomalía del balance entre proteasas y antiproteasas tiene un rol en la hipersensibilidad visceral y en la inflamación de bajo grado.
¿Cuáles son los principales resultados brindados por este estudio?
El objetivo de este estudio era investigar si una perturbación de la microbiota intestinal, asociada a un aumento de las proteasas intestinales, era capaz de inducir una hipersensibilidad visceral. En el estudio se incluyeron niños en lactancia materna de entre 1 y 4 meses de edad. No había diferencias en cuanto a la duración del embarazo, al peso en el nacimiento y a los antecedentes familiares de alergia, entre el grupo con cólicos (n = 7) y el grupo control (n = 7). Por definición (criterios de Roma III), solo la duración de llanto difería, en promedio de 240 ± 95,95 minutos (grupo con cólicos) frente a 24,04 ± 19,65 minutos (grupo control).
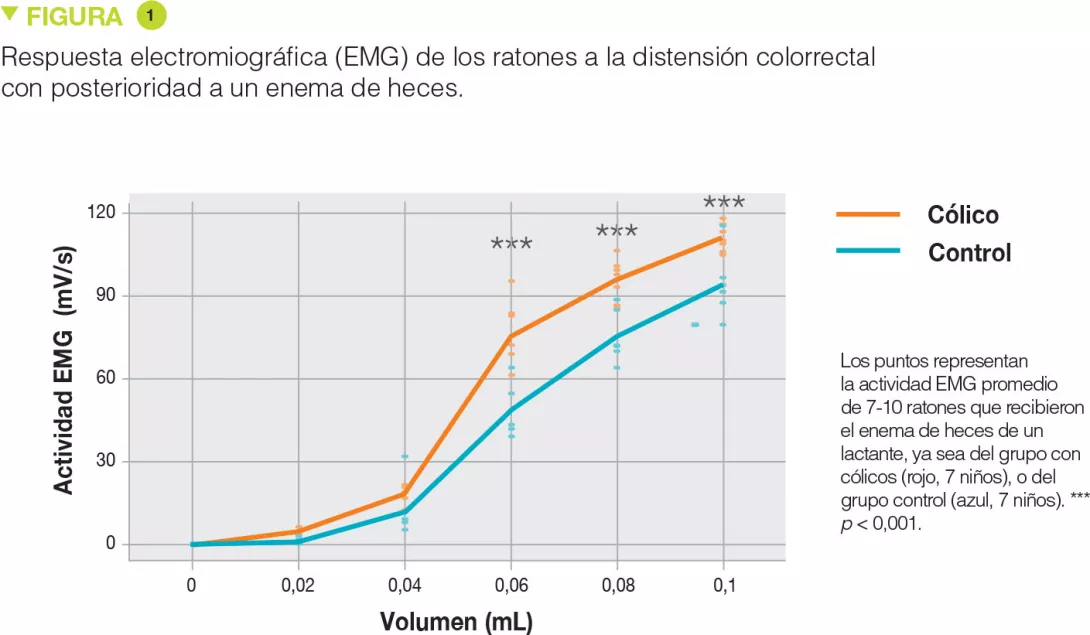
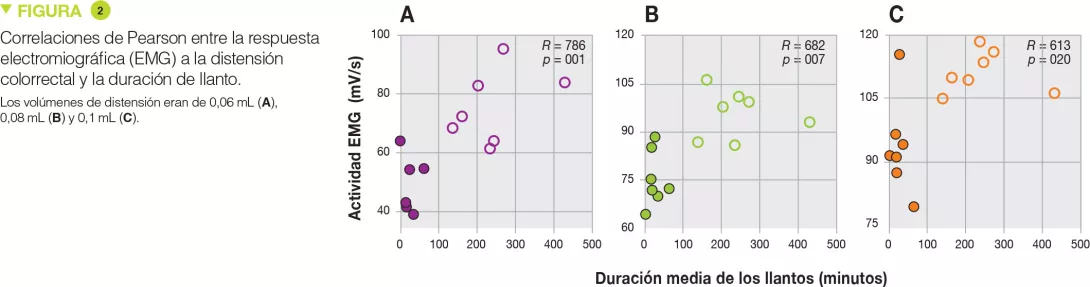
Los enemas rectales de heces de lactantes con cólicos provocaron una hipersensibilidad significativa durante la distensión rectal con volúmenes más importantes (+55 %, p < 0,001 a 0,06 mL; +27 %, p < 0,001 a 0,08 mL y +19 %, p < 0,001 a 0,1 mL) (Figure 1), pero también en respuesta global (aumento del área bajo la curva de +33 %, p < 0,001). Además, existía una correlación positiva con la duración de llanto para esos volúmenes de distensión (Figure 2). Por el contrario, no había diferencias en cuanto a las cantidades de proteasas (serina, y sustancias similares a la tripsina y la elastasa) entre ambos grupos.
Finalmente, el análisis de la microbiota mostró un aumento de la diversidad y una mayor abundancia de Bacteroides vulgatus y de Bilophila wadsworthia en los niños con cólicos de lactante. La abundancia relativa de B. vulgatus estaba asociada positivamente a la hipersensibilidad de los ratones (p = 0,021) y no significativamente a la duración de lo llanto (p = 0,067).
Imagen
¿Cuáles son las consecuencias en la práctica?
Este estudio muestra que un componente contenido en las heces del lactante con cólicos es capaz de desencadenar una hipersensibilidad visceral en ratones. Podría tratarse de un aumento de un compuesto nociceptivo o de la disminución de un compuesto antinociceptivo. No se trata de una alteración del balance proteolítico, pero es posible que ese compuesto sea provocado por una microbiota intestinal diferente. Sin embargo, es necesario realizar estudios complementarios para identificar cuál podría ser ese compuesto, y cuál sería el mecanismo de acción.
Imagen
Summary
Off
Migrated content
Désactivé
Updated content
Désactivé
La mejora de la sensibilidad a la insulina posterior al trasplante de microbiota fecal depende de la composición inicial de la microbiota de los receptore
Artículo comentado - adulto
Por el Pr. Harry Sokol
Servicio de gastroenterología y nutrición, Hospital Saint-Antoine, París, Francia
El comentario del artículo original de Kootte et al. (Cell Metab 2017)
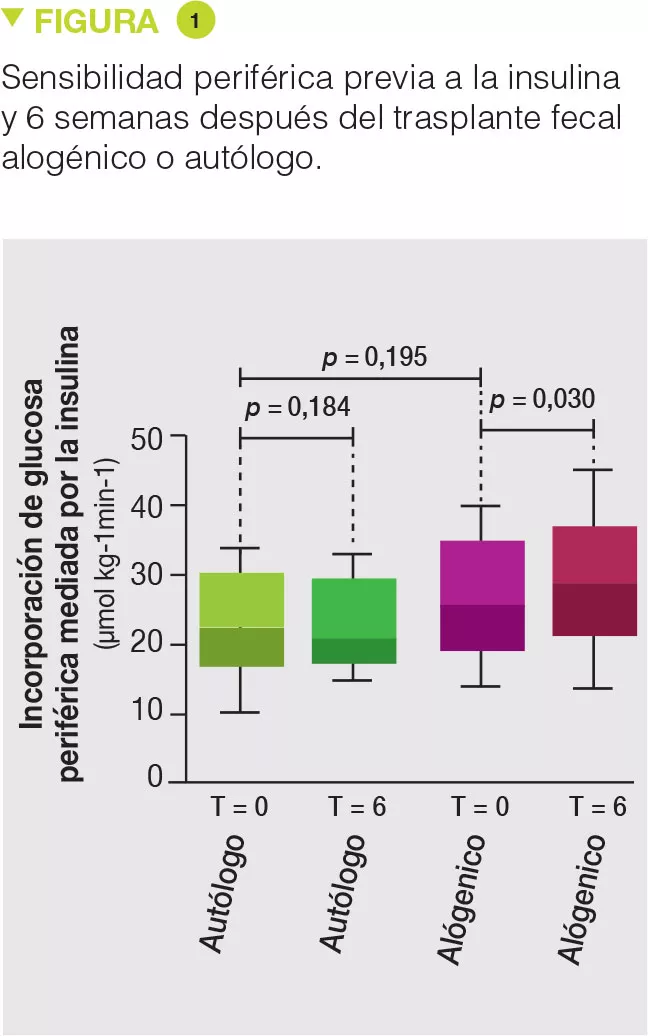
La microbiota intestinal tiene incidencia en la resistencia a la insulina, si bien las pruebas relativas a la relación de causalidad son limitadas. Hemos comparado el efecto de un trasplante de microbiota fecal (TMF) de un donante delgado (alogénico) con el de un autotrasplante (autólogo) en pacientes de sexo masculino con síndrome metabólico. Si bien no se observó ningún cambio metabólico 18 semanas después del TMF, la sensibilidad a la insulina había mejorado significativamente a las 6 semanas en el grupo TMF alogénico, y eso estaba asociado a una modificación de la composición de la microbiota. También hemos informado cambios en las concentraciones de metabolitos plasmáticos como el ácido γ-aminobutírico, y hemos mostrado que la respuesta metabólica posterior al TMF (definida como la mejora de la sensibilidad a la insulina 6 semanas después del TMF) se observa en pacientes con una diversidad microbiana reducida al estado basal. En conclusión, los efectos beneficiosos del TMF de donantes delgados en el metabolismo glucídico se asocian a cambios de la microbiota intestinal y de los metabolitos plasmáticos, y pueden predecirse a partir de la composición basal de la microbiota del receptor.[4]
¿Qué es lo que ya se sabe sobre este tema?
La obesidad y las patologías asociadas a ella, como la diabetes, requieren nuevos enfoques terapéuticos, porque los tratamientos actuales, las modificaciones en el estilo de vida y los tratamientos antidiabéticos no son suficientemente eficaces para reducir la morbilidad y la mortalidad. Durante la última década, los cambios en la composición de la microbiota intestinal han emergido como una nueva estrategia terapéutica potencial para mejorar la sensibilidad a la insulina [1]. Varios estudios han mostrado que la composición de la microbiota intestinal difiere entre animales delgados y obesos, pero también que la composición microbiana podría ser el reflejo de funciones metabólicas alteradas, en particular con una perturbación de los alimentos ingeridos [2]. Finalmente, estos estudios en animales han sugerido una relación causal entre las anomalías de la microbiota y el síndrome metabólico, dado que el fenotipo es transferible por TMF [2]. Si bien numerosos estudios observacionales han sugerido correlaciones entre una composición alterada de la microbiota y el metabolismo humano, ha sido difícil probar la causalidad. Los autores del presente estudio han demostrado previamente, en un pequeño estudio piloto, que el TMF de donante delgado a hombres con síndrome metabólico inducía una mejora del metabolismo glucídico, de forma paralela a cambios en la microbiota fecal y duodenal [3]. Esos resultados animaron a los autores a estudiar los efectos a corto y largo plazo del TMF de donante delgado en la composición de la microbiota intestinal en un grupo más grande de hombres con síndrome metabólico, y a explorar la fisiopatología de la resistencia a la insulina, correlacionando los cambios de la microbiota intestinal con varios marcadores del metabolismo. Además, los autores han intentado identificar las características basales de la microbiota de los receptores que permitan explicar la mejora de la sensibilidad a la insulina en determinados pacientes (identificados como respondedores metabólicos) pero no así en otros (no respondedores).
¿Cuáles son los principales resultados brindados por este estudio?
Treinta y ocho hombres obesos con síndrome metabólico fueron incluidos y aleatorizados en el grupo TMF alogénico (n = 26) o en el grupo autólogo (n = 12). El TMF se realizó por sonda nasoduodenal y se repitió 6 semanas después. Dieciocho semanas después del TMF, no se observaba ningún efecto, ni en la microbiota, ni en los parámetros del síndrome metabólico. Ahora bien, 6 semanas después del TMF, la microbiota del grupo TMF alogénico se había modificado, y los parámetros metabólicos, en especial la sensibilidad a la insulina, habían mejorado, mientras que no se observaba ningún cambio en el grupo TMF autólogo (Figure 1). Contrariamente a lo sucedido en su anterior estudio, no se observó ningún cambio en la concentración de butirato fecal [3]. Por el contrario, el TMF alogénico estaba asociado a un aumento de la concentración de acetato fecal, así como a modificaciones del porcentaje de una treintena de metabolitos sanguíneos, muchos de ellos implicados en el metabolismo del triptófano. En el subgrupo de pacientes que respondieron favorablemente al TMF alogénico, se observaron cambios en la microbiota fecal como, por ejemplo, un aumento de la bacteria Akkermansia muciniphila, cuyos efectos favorables sobre el síndrome metabólico se han demostrado en ratones. Los autores también pusieron en evidencia que la composición basal de la microbiota, así como su poca diversidad, permitían predecir una buena respuesta al TMF.
Imagen
¿Cuáles son las consecuencias en la práctica?
Las intervenciones en la microbiota intestinal, y en particular el TMF, son una pista terapéutica válida en el síndrome metabólico. Sin embargo, existe una fuerte variabilidad interindividual en la respuesta, que puede vincularse tanto a factores del huésped como del donante. Por otra parte, los efectos son relativamente modestos con un único TMF y, en el mejor de los casos, son transitorios. Estrategias más focalizadas, como el uso de probióticos de nueva generación (bacterias provenientes de la microbiota), y con una administración prolongada, son más atractivas y se encuentran actualmente en estudio.
Bibliografia
Summary
Off

Migrated content
Désactivé
Updated content
Désactivé
Tratamiento con antibióticos en niños: consecuencias a corto y largo plazo sobre el microbioma
Síntesis
Por el Pr. Yvan Vandenplas
Por el Prof. Yvan Vandenplas Jefe del Departamento de Pediatría, KidZ Health Castle, Bruselas, Bélgica
Summary
Off

Migrated content
Désactivé
Updated content
Désactivé
Obesidad materna durante el embarazo y tipo de parto
Revisión de prensa
Por el Pr. Ener Cagri Dinleyici
Servicio de Pediatría, Facultad de Medicina, Universidad Eskisehir Osmangazi, Eskisehir, Turquía
Summary
Off
Migrated content
Désactivé
Updated content
Désactivé
La microbiota intestinal modula la respuesta tumoral en pacientes con cáncer
Revisión de prensa
Por el Pr. Ener Cagri Dinleyici
Servicio de Pediatría, Facultad de Medicina, Universidad Eskisehir Osmangazi, Eskisehir, Turquía
Summary
Off
Migrated content
Désactivé
Updated content
Désactivé