Microbiote intestinal #13
Revue de presse
Par le Pr. Markku Voutilainen
Faculté de médecine de l'Université de Turku ; gastro-entérologie, Hôpital universitaire de Turku, Finlande

Microbiote intestinal, régime méditérranéen et maladies cardiovasculaires
Premiers contributeurs à la charge de morbidité dans le monde, les maladies cardiométaboliques telles que les maladies cardiovasculaires (MCV) et le diabète de type 2 (DT2) ont été reliées à la nature individualisée du microbiote intestinal (interactions métaboliques et immunitaires). Même si des études précliniques suggèrent une relation bidirectionnelle entre le microbiote intestinal et le régime alimentaire, il manque toujours des données cliniques solides, notamment concernant le risque de maladie cardiométabolique. L’objectif de cette étude était d’examiner les interactions entre un régime méditerranéen, le microbiote intestinal et le risque de maladie cardiométabolique dans une sous-population de plus de 300 hommes participant à l’étude au long cours Health Professionals Follow-up Study (HPFS). Une interaction significative a été identifiée entre un régime alimentaire sain et le microbiote intestinal vis-à-vis du risque de maladie métabolique. Cette étude montre que l’adhésion à long terme à un régime méditerranéen sain était associée à une variation taxonomique et enzymatique du microbiote intestinal. Le régime alimentaire expliquait 0,7 % de la variation, soit une proportion plus importante que celle induite par le recours aux antibiotiques. L’adhésion au régime méditerranéen était associée à un enrichissement de la dégradation microbienne de fibres alimentaires et de la fermentation en acides gras à chaîne courte induites par les métaboliseurs de fibres anaérobies tels que F. prausnitzii et E. rectale. La faible adhésion au régime méditerranéen, avec la consommation de viande rouge ou transformée, était associée à une augmentation de la synthèse microbienne des acides biliaires secondaires hépatotoxiques principalement induite par C. aerofaciens. Cette étude montre que le régime méditerranéen est capable de réduire le risque de maladie cardiométabolique en l’absence de Prevotella copri : alors qu’une augmentation de l’indice d’adhésion au régime méditerranéen était associée à une diminution du risque d’infarctus du myocarde chez les non-porteurs de Prevotella copri, les porteurs de P. copri présentaient un risque accru. Par conséquent, le profil microbien intestinal pourrait être utilisé pour adapter les interventions alimentaires afin de prévenir les maladies CV. Pour les non-porteurs de P. copri, un régime méditerranéen constituerait la mesure préventive de première ligne, alors que les porteurs de P. copri pourraient davantage bénéficier de l’exercice physique ou des statines pour contrôler le risque CV.
Antibioprophylaxie et antibio-résistance chez les patients leucémiques
Même si l’antibioprophylaxie (AP) peut réduire le risque d’infections graves chez les patients immunodéprimés, elle présente un inconvénient majeur : l’antibiorésistance. La prophylaxie par fluoroquinolone à large spectre peut conduire à la sélection de micro-organismes antibiorésistants et à une résistance croisée aux autres antibiotiques. Dans cette étude, les auteurs ont analysé le résistome gastro-intestinal d’enfants atteints de leucémie aiguë lymphoblastique (LAL) afin de déterminer l’impact de l’AP sur les gènes de résistance aux antibiotiques (GRA). Sur les 49 enfants atteints de LAL, 31 (63 %) ont reçu une prophylaxie par lévofloxacine pendant le traitement d’induction et 18 n’ont pas reçu de prophylaxie. L’association triméthoprime- sulfaméthoxazole a été administrée à titre de prophylaxie contre Pneumocystis jirovecii. Une augmentation de l’abondance relative des gènes de résistance à l’association triméthoprime-sulfaméthoxazole a été détectée dans le microbiote intestinal, augmentation qui n’a pas été modifiée par l’administration prophylactique de lévofloxacine. Les mutations ponctuelles des topo-isomérases des bactéries fécales ont augmenté pendant le traitement chez les patients sous lévofloxacine, mais pas dans le reste de la population de l’étude. La lévofloxacine cible les enzymes topo-isomérases bactériennes qui catalysent des cassures double brin de l’ADN. L’augmentation de la prévalence des gènes de résistance aux fluoroquinolones était limitée et le nombre de patients présentant des mutations des topo-isomérases est resté faible. Même si l’effet sélectif de la lévofloxacine a semblé limité, une augmentation de la fréquence de la résistance aux fluoroquinolones a persisté pendant au moins 2 mois après l’exposition. Par contre, aucune modification n’a été détectée au niveau des gènes de résistance aux aminosides, aux β-lactamines et à la vancomycine ou des gènes de multirésistance après le traitement d’induction, ce qui suggère qu’il n’y a pas de résistance croisée aux autres antibiotiques. En conclusion, la prophylaxie par fluoroquinolone offre une protection à court terme contre les infections mais n’augmente pas le risque de résistance croisée aux autres antibiotiques.
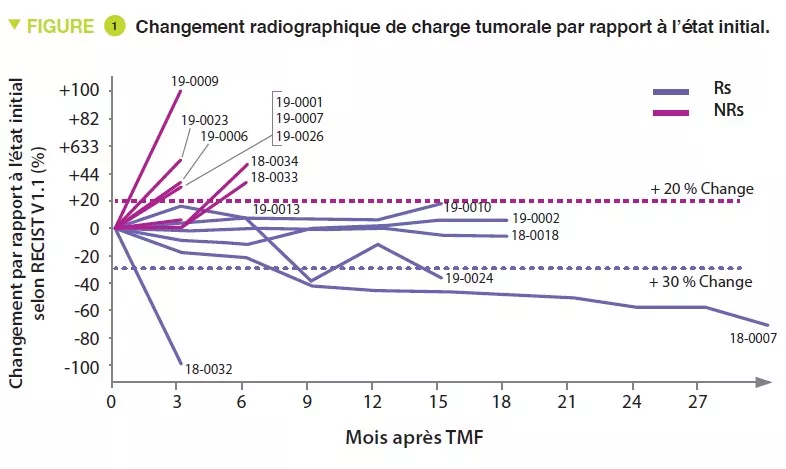
Rôle de la transplantation de microbiote fécal (TMF) dans le traitement du mélanome
L’immunothérapie visant à inhiber la protéine point de contrôle PD-1 (programmed cell death-1) est utilisée chez les patients atteints de mélanome, mais seuls 10 à 20 % obtiennent une rémission complète. Pour augmenter les chances de succès du traitement, la modulation du microbiote intestinal est devenue l’une des pistes les plus prometteuses avec des résultats positifs dans les modèles précliniques. Cependant, elle n’a pas été évaluée dans le cadre d’essais cliniques. Les auteurs ont voulu analyser l’impact de la TMF suivie par une immunothérapie anti-PD-1 sur les cellules immunitaires chez des patients atteints d’un mélanome métastatique réfractaire. La TMF a été réalisée à la fois par coloscopie et par administration orale de gélules fécales et a été suivie de la réintroduction du traitement anti-PD-1. Les selles ont été obtenues auprès de deux donneurs (donneurs 1 et 2) dont le mélanome métastatique avait été traité et qui avaient obtenu une rémission complète. La TMF n’a entraîné aucun événement indésirable modéré ou sévère. Des réponses objectives au traitement du mélanome ont été détectées chez trois patients, tous ayant reçu la TMF du même donneur (1). Un patient a obtenu une rémission complète et deux une rémission partielle. Après la TMF, le microbiote intestinal différait par rapport à la situation initiale chez tous les patients et il était différent en fonction du donneur (1 ou 2). Les répondeurs présentaient une abondance relative plus élevée d’Enterococcaceae, Enterococcus et Streptocccus australis et une abondance plus faible de Veillonella atypica, mais aucune association n’a été détectée entre les taxa microbiens et la réponse au traitement. Après la TMF, on a observé une augmentation de l’expression des gènes liés à la présentation des peptides par les cellules présentant l’antigène (CPA). Les répondeurs présentaient également une augmentation de l’expression des gènes liés à l’activité des CPA, à l’immunité innée et à l’interleukine 12. L’analyse des échantillons tumoraux de tous les receveurs disponibles a révélé une augmentation post-traitement de l’expression de nombreux ensembles de gènes liés à la réponse immunitaire. Cette étude montre que la TMF associée à un anti-PD-1 constitue un traitement sûr et potentiellement efficace dans le mélanome métastatique réfractaire. La modulation du microbiote intestinal pourrait permettre de contrer la résistance à l’immunothérapie.
Microbiote et cancer du sein
Dans cette revue, les auteurs se concentrent sur le microbiote humain tout au long de la vie, sur les liens existant entre le microbiote intestinal/mammaire et le cancer du sein (CS), et sur l’impact de la métabolomique et de la pharmacomicrobiomique sur le risque et le pronostic du CS et les choix thérapeutiques. Les oestrogènes, la forte densité mammaire, le régime alimentaire occidental, l’obésité, l’alcool et des facteurs génétiques sont des facteurs de risque connus de CS, mais la dysbiose du microbiote intestinal joue un rôle clé dans le développement, le traitement et le pronostic du CS par le biais de différents processus biologiques. Les bactéries présentant une activité β-glucuronidase (BGUS) modifient la circulation entéro-hépatique des oestrogènes et peuvent augmenter le risque de CS hormono-dépendant. Comme dans l’intestin, les signatures microbiennes locales du microbiote mammaire chez les patientes atteintes de CS diffèrent de celles des témoins sains. On ignore s’il s’agit là d’une cause ou d’une conséquence, mais il pourrait y avoir un lien entre dysbiose mammaire et CS, influencé par les bactéries et/ou leurs composantes dans le microenvironnement immunitaire local. (sidenote: L’oestrobolome humain Ensemble des gènes bactériens entériques dont les produits sont capables de métaboliser les oestrogènes Plottel CS, Blaser MJ. Microbiome and malignancy. Cell Host Microbe. 2011 Oct 20;10(4):324-35. ) désigne les gènes bactériens entériques dont les produits métabolisent les oestrogènes. L’enzyme BGUS des bactéries intestinales déconjugue les xénobiotiques et les oestrogènes, ce qui conduit à une réabsorption via la circulation entéro-hépatique. Les oestrogènes produits par la BGUS peuvent majorer le risque de CS hormono-dépendant. D’autres bactéries intestinales métabolisent les phyto-oestrogènes qui peuvent conférer une protection contre le CS. Certaines bactéries intestinales produisent de l’équol et des entérolignanes qui peuvent réduire le risque de CS hormono- dépendant. 20 à 30 % de la population occidentale possède des micro-organismes (famille des Coriobacteriaceae) convertissant l’isoflavone en équol qui présente une affinité pour les récepteurs aux oestrogènes, et une activité antiandrogénique et antioxydante. Les récepteurs des acides gras libres au niveau de l’intestin sont activés par les acides gras à chaîne courte et pourraient participer à la suppression tumorale. Les microbiotes mammaire et intestinal peuvent moduler le microenvironnement du CS par différents mécanismes : activation d’une prolifération épithéliale aberrante, sécrétion de facteurs de croissance, mutations génomiques, perturbations du microenvironnement métabolique local et angiogenèse. Les bactéries intestinales peuvent par exemple inactiver la doxorubicine et la gemcitabine. Le microbiote intestinal a même un rôle double dans l’efficacité de la radiothérapie, avec des effets bénéfiques et protecteurs ou délétères et résistants. En conclusion, chez les patientes atteintes d’un CS, le microbiote pourrait constituer un facteur pronostique et prédictif de la réponse au traitement. À l’avenir, la modulation du microbiote pourrait améliorer le devenir des patientes atteintes d’un CS.