Une nouvelle étude révèle que la transplantation de microbiote fécal améliore les symptômes du syndrome du côlon irritable et la qualité de vie des patients, et cela même un an après le traitement, mais qu'il est essentiel d'avoir un donneur en bonne santé.
Douleurs, crampes, ballonnements, diarrhées, constipations... Le syndrome du côlon irritable est une maladie qui se manifeste par un ensemble de symptômes abdominaux qui vont et viennent au cours de la vie du patient. Ces symptômes peuvent être exacerbés par le stress, les changements émotionnels ou certains aliments… et altérés considérablement la qualité de vie du patient. S’il n’existe pas de problème anatomique ou structurel de l'intestin des personnes souffrant de ce syndrome, le microbiote intestinal, en revanche, est souvent mis en cause.
Un donneur unique avec un super microbiote ?
Les chercheurs de cette étude ont testé dans un essai clinique l'efficacité d’une transplantation de microbiote fécal en utilisant des échantillons de selles d'un seul homme caucasien âgé de 36 ans1 qui cochait toutes les cases du « super donneur » : en bonne santé, IMC normal, faisant régulièrement de l'exercice, né par voie vaginale et ayant été allaité. Mieux, il ne prenait aucun médicament, n'avait reçu que trois traitements antibiotiques au cours de sa vie et prenait régulièrement des compléments alimentaires. Dans cet essai clinique, la transplantation de microbiote fécal s'est avérée efficace chez des patients souffrant du syndrome du côlon irritable. Cependant, ces résultats n'ont été observés que 3 mois après la transplantation, et plusieurs questions restent à résoudre, notamment celle de savoir si l'effet clinique de la transplantation se maintient à long terme. Lors d’une présente étude, les chercheurs ont continué le suivi de ces patients pendant un an.
Des bénéfices toujours présents après 1 an
La plupart des patients qui avaient répondu à la transplantation de microbiote fécal après 3 mois ont maintenu leur réponse après 1 an. Autre résultat encourageant, leurs symptômes abdominaux, leur fatigue et leur qualité de vie s'étaient nettement améliorés par rapport à trois mois après la transplantation. Encore mieux, entre 32 et 45% des patients, selon les groupes, ont connus une rémission complète au cours de l’année de suivi. L’analyse complète du microbiote intestinal des patient a montré des changements dans le profil bactérien intestinal et une diminution significative de l’indice de dysbiose.
En conclusion, la transplantation de microbiote fécal à partir d’un « super-donneur » permettrait de rétablir le microbiote intestinal et diminuer les symptômes des patients souffrants du syndrome du côlon irritable.
La dysbiose est une rupture du délicat équilibre entre les milliards de micro-organismes du microbiote humain et de ses bonnes relations avec notre corps. Génétique, alimentation déséquilibrée, antibiotiques… les facteurs de dysbiose sont divers et la plupart du temps, multiples. Aujourd’hui, la recherche scientifique montre qu’une dysbiose du microbiote intestinal, la plus étudiée, mais aussi de nos autres microbiotes comme celui du système vaginal, cutané ou du poumon, est associée à différentes maladies, du syndrome du côlon irritable à des troubles métaboliques comme l’obésité, ou même la sinusite chronique et l’eczéma. Comment le microbiote peut-il se déséquilibrer ? Quelles sont les conséquences d’une dysbiose sur notre santé ? Comment le microbiote retrouve-t-il son équilibre ?
Tout d'abord revenons sur le mot « dysbiose » : l’étymologie de ce terme scientifique est très simple ! En grec, le mot bios signifie « vivant » et le préfixe dys- signifie « mauvais ».
Une « dysbiose » peut être définie comme une altération dans la composition et la fonction du microbiote. Cette altération résulte d’une combinaison de facteurs environnementaux et de facteurs spécifiques à chaque personne1.
Étant donné que les micro-organismes colonisent tout notre corps, il est possible d’observer une dysbiose dans les microbiotes suivants :
Microbiote intestinal : différentes maladies ont été associées à la dysbiose intestinale (diarrhée post-antibiotiques14, gastro-entérite17, colique du nourrisson...44).
Microbiote cutané : la dysbiose est souvent associée à des pathologies (acné45, dermatite atopique46).
Microbiote vaginal : une dysbiose vaginale est associée à une vaginose bactérienne1, une candidose47, une diminution de la fécondité48 ou une augmentation du risque d’accouchement prematuré1.
Microbiote ORL (oreille, nez, gorge) : diverses maladies peuvent être associées à un déséquilibre du microbiote oral, auriculaire ou nasopharyngé.
Microbiote pulmonaire : la dysbiose peut contribuer au développement d’infections respiratoires hivernales49, de l’asthme50 et de la mucoviscidose51.
Microbiote urinaire : les études publiées à ce jour ont montré que le microbiote urinaire peut jouer un rôle dans les infections urinaires52.
Une dysbiose sur le grill : le microbiote intestinal
Le microbiote intestinal est le principal microbiotedu corps humain 2. Il abrite au moins 1000 espèces différentes3 de microorganismes : des bactéries, mais aussi des champignons, des virus… Les groupes des Firmicutes (qui comprennent les lactobacilles, ces « bonnes bactéries » bien connues) et des Bacteroidetes représentent à eux deux pour 70% à 90% de la communauté bactérienne de notre intestin 2,4. Notre microbiote contient également des Actinobacteria, auxquelles appartiennent les bifidobactéries, réputées pour leurs bienfaits. D’autres micro-organismes de notre microbiote pourraient nous rendre malades, ils sont dits potentiellement (sidenote:
pathogènes
Un pathogène est un microorganisme qui cause, ou peut causer, une maladie
Pirofski LA, Casadevall A. Q and A: What is a pathogen? A question that begs the point. BMC Biol. 2012 Jan 31;10:6.), mais sont minoritaires2. Une dysbiose se traduit par un ou plusieurs de ces phénomènes :
Les proportions entre ces grandes familles bactériennes se modifient de façon importante, avec notamment une perte de lactobacilles et de bifidobactéries5;
Des micro-organismes utiles qui vivent normalement dans notre microbiote (dits « commensaux ») diminuent ou disparaissent1 ;
La diversité des micro-organismes présents dans le microbiote s’appauvrit : il y a moins d’espèces différentes5;
Les micro-organismes potentiellement pathogènes du microbiote prolifèrent1,5.
Conséquences : notre microbiote se fragilise et les « mauvaises » bactéries prennent le dessus sur les « bonnes »2. Il protège moins facilement notre organisme des agressions et assure moins efficacement ses rôles essentiels pour notre forme et notre santé1,6.
1000
Il abrite au moins 1000 espèces différentes de microorganismes.
Qu’est-ce que la dysbiose ?
Bien que la dysbiose ne soit pas considérée comme une maladie, elle est associée à différents problèmes de santé et peut contribuer au développement ou à l’exacerbation de certaines pathologies.
Un déséquilibre du microbiote propre à chacun
Cependant, la dysbiose n’est pas un terme universel qui peut s’appliquer à tous et dans toutes les circonstances1! En effet, influencée par nos gènes et les micro-organismes (déf : organismes vivants qui sont trop petits pour être vus à l'œil nu. Ils incluent les bactéries, les virus, les champignons, les archées, les protozoaires, etc… et sont communément appelés " (sidenote: https://microbiologysociety.org/why-microbiology-matters/what-is-microbiology.html)") qui ont colonisé notre corps dans nos premières années de vie, la composition de notre microbiote est personnelle. Elle varie si considérablement entre les individus qu’elle pourrait être aussi propre à chacun qu’une empreinte digitale7. Mais elle peut aussi évoluer selon notre âge, notre état de santé, la survenue d’un stress, notre alimentation, notre lieu de vie, les médicaments que nous prenons…8. Ainsi, chacun de nous peut avoir « sa » dysbiose quand notre microbiote se déséquilibre et ne fonctionne plus correctement au sein de notre organisme1.
Mais alors c’est quoi un microbiote équilibré ?
Le préfixe dys- de dysbiose s’oppose à eu- (« bon ») ou à sym- (« avec »). On parle ainsi d’« eubiose » ou de « symbiose » quand notre microbiote est en bonne santé : il interagit en harmonie avec notre organisme et sa communauté microbienne est équilibrée1.
En effet, il existe un bénéfice mutuel entre les milliards de micro-organismes qui peuplent notre microbiote et notre corps9. Chacun y trouve son compte : l’organisme fournit « le gîte et le couvert » aux micro-organismes du microbiote. En retour, ceux-ci contribuent à de nombreuses fonctions importantes de notre corps, comme la digestion, l’assimilation des nutriments, la protection de l’imperméabilité de la paroi intestinale et la lutte contre les germes indésirables2,8,10. C’est un véritable travail d’équipe !
Les différents micro-organismes de la communauté de notre microbiote, y compris ceux qui pourraient être pathogènes, sont présents en nombre et en proportions adéquates pour cohabiter pacifiquement et assurer leurs fonctions bénéfiques à l’organisme. Mais l’équilibre subtil entre les écosystèmes microbiens de notre corps peut se rompre : l’eubiose se transforme alors en dysbiose8.
Qu’est ce qui provoque une dysbiose ?
Comme l’indique sa définition, une dysbiose apparaît sous l’influence de nombreux facteurs très différents, souvent intriqués5. On peut toutefois distinguer :
Ceux qui sont liés à l’individu lui-même, comme :
la génétique1;
l’âge11;
certaines pathologies et blessures1 ;
Ceux qui sont liés à son environnement, comme :
la prise de médicaments : antibiotiques, anti-inflammatoires…2,5,
les infections12;
le mode de vie : alimentation déséquilibrée ou changements d’alimentation, stress, tabagisme, hygiène inadaptée…1, 5,8 ;
Les antibiotiques : irremplaçables, mais perturbateurs du microbiote
Les antibiotiques ont représenté l’un des progrès thérapeutiques les plus importants du XXème siècle. Depuis la découverte de la pénicilline en 1928, ils ont sauvé des millions de vie13. Cependant, en détruisant les germes néfastes mais aussi les « bonnes » bactéries, ils déséquilibrent le microbiote. A court terme, la dysbiose engendrée par les antibiotiques peut se traduire par une diarrhée14 ou une mycose vaginale15. La dysbiose intestinale induite par les antibiotiques est également soupçonnée d’avoir un impact à long terme, notamment quand les antibiotiques sont pris pendant l’enfance, en augmentant le risque de différentes maladies chroniques comme l’obésité ou les allergies…
Les infections : quand les microbes attaquent !
Lors des infections comme les gastro-entérites virales ou les intoxications alimentaires par des salmonelles, des germes néfastes et offensifs envahissent le microbiote. Ils ne proviennent pas du microbiote humain, mais de l’extérieur, transmis par exemple par des mains ou des aliments contaminés. Ces infections provoquent une forte réaction de notre système immunitaire, une inflammation au niveau de l’intestin et des diarrhées. Tout ceci aboutit à des perturbations abruptes de l’équilibre de notre flore intestinale. De plus, les microbes en cause dans ces infections peuvent également favoriser le développement d’autres bactéries potentiellement pathogènes déjà présentes dans le microbiote. Les infections provoquent donc des dysbioses, dont profitent tous les germes néfastes1,12,17,18!
L’alimentation : l’équilibre du microbiote dans nos assiettes
Ce que nous mangeons a un impact sur notre microbiote tout au long de notre vie. Un changement brutal d’alimentation, dans sa composition ou sa quantité, peut déclencher une dysbiose. Mais pas seulement : si les variations normales de nos menus d’un jour à l’autre n’entraînent que des modifications transitoires du microbiote, notre type d’alimentation peut modifier durablement l’écosystème digestif5 et peut, à terme, être facteur de dysbiose. Ainsi, des études suggèrent qu’une alimentation « à l’occidentale » riche en graisses, en sucres et en protéines favoriseraient les déséquilibres du microbiote intestinal, tandis qu’une alimentation variée et riche en fruits et légumes pourrait l’en protéger1,19.
Les antibiotiques
Ils ont sauvé des millions de vies, mais leur usage excessif et inapproprié pose aujourd’hui de sérieux enjeux de santé, notamment en raison de la résistance aux antimicrobiens et de la dysbiose du microbiote. Chaque année, l’OMS organise la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens (WAAW) dans le but de mieux faire connaître le phénomène de résistance aux antimicrobiens. Consultez cette page dédiée pour en savoir plus.
La résistance aux antimicrobiens survient lorsque les bactéries, les virus, les parasites et les champignons évoluent et résistent aux effets des médicaments. En raison de cette pharmacorésistance, les antibiotiques et autres médicaments antimicrobiens perdent leur efficacité et les infections deviennent plus difficiles, voire impossibles à traiter, ce qui augmente le risque de propagation des maladies, de pathologie grave et de décès.
Organisée du 18 au 24 novembre, cette campagne invite le grand public, les professionnels de santé et les décideurs politiques à faire un usage raisonné des antibiotiques, des antiviraux, des antifongiques et des antiparasitaires afin de prévenir le développement accru de la résistance aux antimicrobiens. "
A chaque microbiote, sa dysbiose et ses pathologies
La dysbiose : cause ou conséquence d’une maladie ?
De nombreuses études comparant les microbiotes de personnes saines et malades ont montré qu’une dysbiose était associée à diverses pathologies chroniques : des maladies intestinales comme le syndrome du côlon irritable ou la maladie de Crohn, mais aussi l’obésité, les allergies, l’asthme et certains cancers...1 Mais est-ce la dysbiose qui provoque la maladie ou la maladie qui provoque la dysbiose ? Pour les scientifiques, la réponse n’est pas toujours évidente mais fait actuellement l’objet de nombreuses recherches.
Pour y voir plus clair, des chercheurs ont lancé en 2019 Homo symbiosus, un grand projet de recherche qui vise à mieux déterminer pourquoi et comment tant de maladies chroniques sont associées à une dysbiose intestinale. Les chercheurs ont émis l’hypothèse que tous « ces phénomènes de dysbiose intestinale, de prolifération microbienne, d’inflammation et de fragilisation de la paroi intestinale » s’entretiennent mutuellement10.
Une dysbiose du microbiote intestinal est impliquée dans des maladies variées : digestives, mais aussi métaboliques22, allergiques23 et même mentales24. Mais le corps humain abrite également des écosystèmes microbiens spécifiques au niveau de la peau25, des voies urinaires26, dans le vagin27 ou encore dans la bouche28 et nos poumons29… dont la composition peut se déséquilibrer et être associée à des maladies spécifiques.
Comment le microbiote retrouve-t-il son équilibre ?
Normalement, après un épisode de dysbiose, le microbiote est capable de retrouver naturellement son équilibre initial (bien qu’on ne retrouve jamais totalement la composition de départ) : on dit qu’il est « résilient » 30. Mais parfois, cette « re-biose », autrement dit le retour à l’équilibre microbien, peut prendre du temps : par exemple, même chez un adulte en bonne santé, elle peut prendre six mois après la prise d’un antibiotique 31. Il arrive enfin que la dysbiose entraîne un état de déséquilibre qui va se prolonger et s’auto-entretenir durablement sans réussir à se rétablir complètement, ce qui peut être dommageable pour la santé 1.
Face à une dysbiose, que faire ? Plusieurs solutions s’offrent à nous pour rétablir l’équilibre d’un microbiote et retrouver une meilleure santé.
Les probiotiques : des micro-organismes bénéfiques en renfort
Les probiotiques sont « des micro-organismes vivants qui, lorsqu'ils sont administrés en quantités adéquates, confèrent un bénéfice pour la santé de l'hôte » 32,33. Vous trouverez ici une page dédiée aux probiotiques à leur fonctionnement, leur fabrication, comment bien les choisir... Retrouvez notre page consacrée aux probiotiques.
Les prébiotiques, pour nourrir notre microbiote
Principalement issus de fibres alimentaires (fructo-oligosaccharides, galacto- oligosaccharides, inuline…), les prébiotiques sont des substrats ou éléments nutritifs non digestibles, utilisés par les micro-organismes du microbiote et qui ont des effets favorables sur la santé34,35. Vous trouverez ici plus d’informations sur leur façon d’agir sur le microbiote. Les produits spécifiques associant des probiotiques et des prébiotiques sont appelés symbiotiques36,37.
Une alimentation saine, pour préserver son bon fonctionnement
Ce que nous mangeons, aussi bien la qualité que la diversité de nos aliments, contribuent à l’équilibre de notre microbiote intestinal 38,39, mais aussi va influencer sa composition et par ricochet être responsable de provoquer certaines maladies22. N’hésitez pas à demander conseil à votre médecin généraliste et /ou diététicien pour mieux connaître les aliments qui ont des effets bénéfiques ou nocifs afin de garder vos intestins en pleine forme40 et de rester en bonne santé !
La transplantation de microbiote une technique d’espoir
Comme les autres organes, le microbiote peut être greffé sur un autre individu afin d’essayer de rétablir l’équilibre de son écosystème microbien 41,42. Pour le moment, cette approche thérapeutique est bien documentée pour le microbiote intestinal, connue sous le nom de transplantation de microbiote fécal (TMF), elle n’est autorisée cependant que pour le traitement des infections récurrentes à Clostridioides difficile41. Elle fait l’objet de recherches intensives pour d’autres pathologies intestinales41… Pour la sphère vaginale, la transplantation de microbiote vaginal (TMV) est à l’essai et pourrait représenter une option prometteuse pour le traitement de la vaginose bactérienne réfractaire ou récurrente.43 Les études sur la transplantation de microbiote cutané sont encore rare, mais les premiers résultats sont prometteurs45,46.
Une formidable avancée médicale victime de son succès. Depuis leur découverte au début du XXe siècle, les antibiotiques ont sauvé des millions de vies. Mais l’utilisation massive et parfois inappropriée de ces traitements les rend de plus en plus inefficaces pour traiter les infections. Résultats : aujourd’hui, de nombreuses bactéries font de la résistance ! Mais l’usage excessif ou inadapté n’est pas le seul responsable de l’antibiorésistance.
Une étude1révèle queles voyages internationaux favorisent l’acquisition de gènes de résistance aux antibiotiques et pourraient contribuer à la propagation de l’antibiorésistance. Embarquement immédiat pour quelques explications.
Comme chaque année depuis 2015, la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens (WAAW) nous sensibilise à l’accroissement de la résistance aux (sidenote:
Antibiorésistance
On parle de résistance aux antimicrobiens lorsque les bactéries, les virus, les champignons et les parasites ne répondent plus aux médicaments du fait de leur évolution dans le temps. Les antibiotiques et autres médicaments antimicrobiens perdent leur efficacité et les infections deviennent de plus en plus difficiles voire impossibles à traiter. Cette résistance aux microbiens augmente le risque de propagation, de forme grave de la maladie voire de décès. L’antibiorésistance correspond à la résistance des antibiotiques aux bactéries.
Source : Résistance aux antimicrobiens. 26 Octobre 2020.
) (également appelée antibiorésistance). Ce phénomène, lié au mauvais usage ou à une consommation excessive d’antibiotiques, désigne la capacité d’une bactérie à résister à l’action d’un antibiotique. Depuis 2020, la WAAW a élargi la portée de sa campagne aux (sidenote:
Antimicrobiens
Médicaments – comme les antibiotiques, les antiviraux, les antifongiques et les antiparasitaires – utilisés pour prévenir et traiter les infections chez les êtres humains, les animaux et les végétaux.
WHO Antimicrobial Resistance; Nov 2023) : antiviraux, antifongiques, antiparasitaires…c’est à dire des médicaments indispensables pour lutter contre les (sidenote:
Microorganismes
Organismes vivants qui sont trop petits pour être vus à l'oeil nu. Ils incluent les bactéries, les virus, les champignons, les archées, les protozoaires, etc… et sont communément appelés "microbes".
Source : What is microbiology? Microbiology Society. ) (sidenote:
pathogènes
Un pathogène est un microorganisme qui cause, ou peut causer, une maladie
Pirofski LA, Casadevall A. Q and A: What is a pathogen? A question that begs the point. BMC Biol. 2012 Jan 31;10:6.) - En réduisant les possibilités de traitement des infections, l’antibiorésistance menace notre santé à tous2. De nombreuses recherches s’attachent donc à comprendre son expansion pour mieux la contrôler voire la stopper.
Les antibiotiques sauvent des vies ! Saviez-vous qu’ils ont aussi un impact sur votre microbiote ? Saviez-vous que l’usage abusif ou excessif des antibiotiques peut engendrer une résistance aux antimicrobiens ? Connaissez-vous la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens (WAAW) ? Découvrez toutes les informations sur cette page dédiée:
Nous savons aujourd’hui que l’antibiorésistance est principalement due à l’utilisation excessive des antibiotiques en médecine humaine, mais aussi dans l’élevage et l’agriculture2. Une récente étude nous révèle toutefois un mécanisme de propagation inattendu : nos vacances et nos déplacements professionnels dans des pays exotiques ! En effet, les voyages internationaux favoriseraient la propagation des (sidenote:
Gène
Un gène est l'unité physique et fonctionnelle de base de l'hérédité. Les gènes sont constitués d'ADN.
Source : What is a gene?MedlinePlus.gov. ) de résistance aux antimicrobiens au niveau intestinal. Des chercheurs ont réuni 190 voyageurs néerlandais, répartis-en 4 sous-groupes en fonction de leur destination. Les lieux de séjour ont été choisis dans des zones fortement touchées par l’antibiorésistance : Asie du Sud-Est, Asie du Sud, Afrique du Nord et Afrique de l’Est. Ces chercheurs ont cherché à déterminer si les déplacements internationaux vers ces régions pouvaient faciliter leur dissémination vers des régions plus épargnées. Ainsi, pour évaluer le portage de ces gènes au niveau intestinal, un échantillon de selles a donc été prélevé chez chaque participant avant et après le voyage.
Image
Une valise diplomatique de l’antibiorésistance au niveau intestinal ?
Grâce à l’utilisation d’une technique de pointe (la (sidenote:
Métagénomique
Méthode d'étude du contenu génétique d'échantillons issus d'environnements complexes (intestin, océan, sols, air, etc.) prélevés dans la nature (par opposition à des échantillons cultivés en laboratoire). Cette approche permet une description des gènes qui sont contenus dans l'échantillon, mais aussi un aperçu du potentiel fonctionnel d'un environnement.
Source : Riesenfeld CS, Schloss PD, Handelsman J. Metagenomics: genomic analysis of microbial communities. Annu Rev Genet. 2004;38:525-52. )), l’équipe a ainsi constaté une hausse du nombre de gènes de résistance aux antibiotiques entre le départ et le retour, en particulier chez les voyageurs revenant d’Asie du Sud-Est. Au total, une cinquantaine de gènes de résistance aux antibiotiques ont été détectés au cours des voyages. Parmi eux, des gènes de résistances classiques et bien connus aux antibiotiques (dont la famille des β-lactamines, des tétracyclines fluoroquinolones entre autres), mais aussi des nouveaux gènes jamais identifiés auparavant.
Voyager : un enjeu de santé publique ?
Les résultats de cette étude sont sans appel : les voyageurs internationaux, colonisés par des gènes de résistance au cours de leurs voyages, pourraient à leur insu rapporter dans leur valise des bactéries résistances aux antibiotiques. Face au risque de propagation, les auteurs tirent la sonnette d’alarme et soulignent l’importance d’engager rapidement des actions dans les pays particulièrement touchés par l’antibiorésistance. Un appel qui fait écho avec la campagne de l’OMS.
Qu'est-ce que la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens ?
La résistance aux antimicrobiens survient lorsque les bactéries, les virus, les parasites et les champignons évoluent et résistent aux effets des médicaments. En raison de cette pharmacorésistance, les antibiotiques et autres médicaments antimicrobiens perdent leur efficacité et les infections deviennent plus difficiles, voire impossibles à traiter, ce qui augmente le risque de propagation des maladies, de pathologie grave et de décès.
Organisée du 18 au 24 novembre, cette campagne invite le grand public, les professionnels de santé et les décideurs politiques à faire un usage raisonné des antibiotiques, des antiviraux, des antifongiques et des antiparasitaires afin de prévenir le développement accru de la résistance aux antimicrobiens.
Depuis la découverte de la pénicilline en 1928, l’utilisation généralisée des antibiotiques a permis de sauver plusieurs millions de vies. Principale arme dans la lutte contre les infections bactériennes, les antibiotiques ont permis de gagner près de 20 ans d’espérance de vie parallèlement aux vaccinations1.
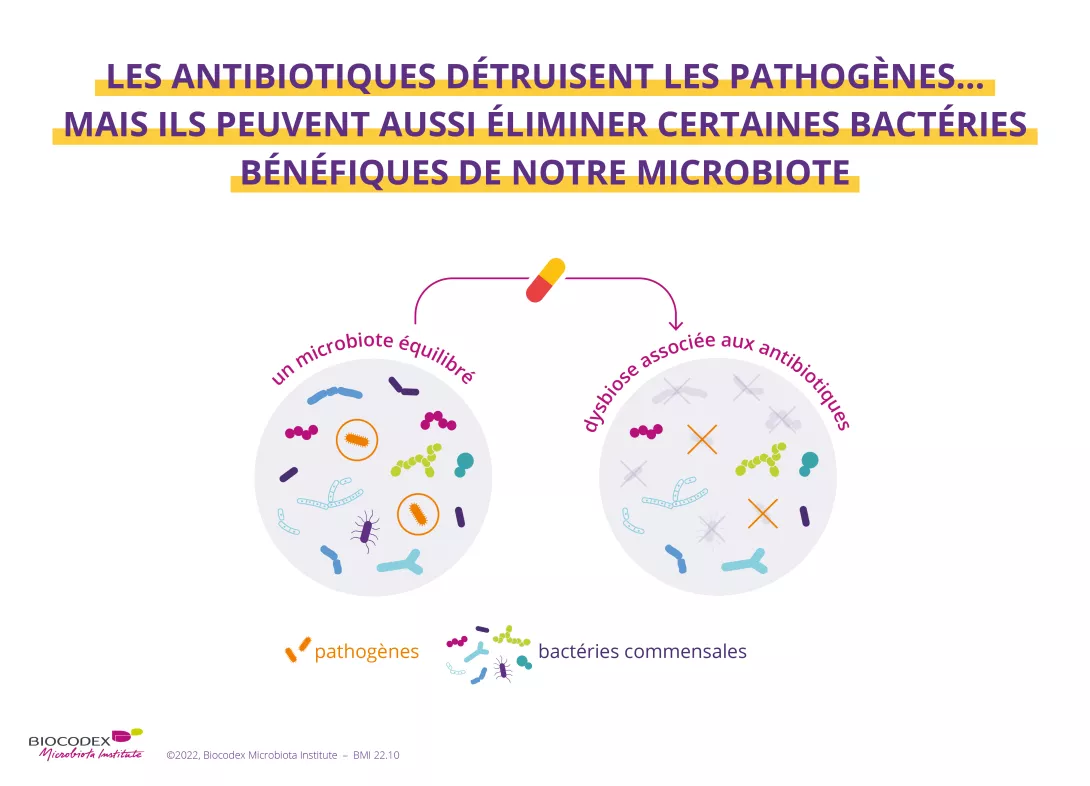
2. Les antibiotiques détruisent les espèces responsables des infections mais aussi les bonnes bactéries
Intestin, vagin, poumon, peau… Plusieurs parties de notre corps abritent des micro-organismes (bactéries, champignons, virus). On les appelle les microbiotes2. Les antibiotiques, s'ils éradiquent les germes pathogènes responsables de notre infection, peuvent également détruire certaines bactéries bénéfiques au sein de notre microbiote et provoquer un déséquilibre plus ou moins important au sein de cet écosystème. C’est ce que l’on appelle une (sidenote:
Dysbiose
La « dysbiose » n’est pas un phénomène homogène : elle varie en fonction de l’état de santé de chaque individu. Elle est généralement définie comme une altération de la composition et du fonctionnement du microbiote, provoquée par un ensemble de facteurs environnementaux et liés à l’individu, qui perturbent l’écosystème microbien.
Levy M, Kolodziejczyk AA, Thaiss CA, et al. Dysbiosis and the immune system. Nat Rev Immunol. 2017;17(4):219-232.)3. Tous les microbiotes du corps sont concernés : le microbiote intestinal mais également le microbiote cutané4, pulmonaire5, ORL6, urinaire7 et vaginal8.
3. Les antibiotiques peuvent engendrer des effets secondaires
En induisant une dysbiose, les antibiotiques peuvent entrainer des conséquences néfastes pour la santé. La principale complication à court terme est la modification du transit chez certains patients. Celle-ci se traduit le plus souvent par une diarrhée, le microbiote intestinal étant moins apte à remplir ses fonctions protectrices. Cette diarrhée associée aux antibiotiques est généralement d'intensité légère à modérée et son incidence varie selon l’âge, le type d’antibiotiques, le contexte… Elle peut toucher jusqu’à 35%9,10,11 des patients et chez les enfants, ce pourcentage peut atteindre jusqu'à 80%9. Dans 10 à 20% des cas, la diarrhée résulte d'une infection par Clostridioides difficile (C. difficile)11 : cette bactérie colonise le microbiote intestinal et va devenir pathogène sous l’influence de certains facteurs (une prise d’antibiotiques par exemple). Les conséquences cliniques sont variables, allant d’une diarrhée modérée à des symptômes beaucoup plus graves, voire la mort11.
35%
Elle peut toucher jusqu’à 35% des patients
80%
jusqu’à 80 % chez les enfants
4. Les antibiotiques peuvent engendrer des effets secondaires à plus long terme
La diarrhée associée aux antibiotiques n’est pas la seule manifestation de la dysbiose associée aux antibiotiques. Cette dernière serait responsable d’effets à plus long terme lorsqu’elle survient très tôt dans la vie. En effet, la période périnatale, qui se caractérise par le développement du microbiote intestinal et la maturation du système immunitaire, constitue une période particulièrement sensible12 : la dysbiose induite par la prise d’antibiotiques durant cette phase semble être un facteur de risque dans la survenue de maladies chroniques (obésité, diabète, asthme, maladies inflammatoires chroniques de l’intestin)13.
5. L’usage inadapté des antibiotiques est responsable de la résistance aux antibiotiques
La résistance aux antibiotiques - ou antibiorésistance – correspond au fait qu’un traitement antibiotique ne soit plus efficace sur une infection bactérienne1. A quoi est-ce dû ? Les antibiotiques ne sont efficaces que sur les bactéries, et n’ont aucune action sur les virus (la grippe par exemple)14. Leur utilisation inadaptée (en cas d’infection virale par exemple) ou excessive - chez l’homme ou l’animal - accélère ce phénomène. La résistance aux antibiotiques entraine des hospitalisations plus longues, une hausse des dépenses de santé et une augmentation des décès. C’est pourquoi celle-ci constitue, à l’échelle mondiale, un enjeu majeur de santé publique1.
6. Il existe, chaque année, une Semaine mondiale pour favoriser le bon usage des antibiotiques
Chaque année, du 18 au 24 novembre, l’OMS organise la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens qui vise à mieux faire connaître le phénomène de résistance aux (sidenote:
Antimicrobiens
Catégories de médicaments regroupant les antibiotiques (action contre les bactéries), antiviraux (contre les virus), antiparasitaires (contre les parasites) et antifongiques (contre les champignons)
WHO Antimicrobial Resistance; Oct 2020) et d’encourager le grand public, les professionnels de santé et les responsables politiques à adopter de meilleures pratiques pour lutter contre l’émergence et la propagation des résistances. En tant qu’expert des microbiotes, le Biocodex Microbiota Institute se joint à cette initiative.
Cette campagne, qui se tiendra du 18 au 24 novembre, encourage le grand public, les professionnels de santé et les décideurs à faire un bon usage des antimicrobiens afin d'éviter l'apparition de résistance.
Côté pile, ils demeurent un pilier de notre arsenal thérapeutique en sauvant des millions de vies chaque année. Côté face, ils pertubent notre microbiote et peuvent avoir de lourdes conséquences sur notre santé. Passage en revue des 6 points essentiels pour les utiliser à bon escient.
Depuis la découverte de la pénicilline en 1928, l’utilisation généralisée des antibiotiques a permis de sauver plusieurs millions de vies. Principale arme dans la lutte contre les infections bactériennes, les antibiotiques ont permis de gagner près de 20 ans d’espérance de vie parallèlement aux vaccinations1.
Image
2. Les antibiotiques détruisent les espèces responsables des infections mais aussi les bonnes bactéries
Intestin, vagin, poumon, peau… Plusieurs parties de notre corps abritent des (sidenote:
Micro-organismes
Organismes vivants qui sont trop petits pour être vus à l'œil nu. Ils incluent les bactéries, les virus, les champignons, les archées, les protozoaires, etc… et sont communément appelés "microbes".
What is microbiology? Microbiology Society.) (bactéries, champignons, virus). On les appelle les microbiotes2. Les antibiotiques, s'ils éradiquent les germes pathogènes responsables de notre infection, peuvent également détruire certaines bactéries bénéfiques au sein de notre microbiote et provoquer un déséquilibre plus ou moins important au sein de cet écosystème. C’est ce que l’on appelle une (sidenote:
Dysbiose
La « dysbiose » n’est pas un phénomène homogène : elle varie en fonction de l’état de santé de chaque individu. Elle est généralement définie comme une altération de la composition et du fonctionnement du microbiote, provoquée par un ensemble de facteurs environnementaux et liés à l’individu, qui perturbent l’écosystème microbien.
Levy M, Kolodziejczyk AA, Thaiss CA, et al. Dysbiosis and the immune system. Nat Rev Immunol. 2017;17(4):219-232.)3.
3. Les antibiotiques peuvent engendrer des effets secondaires
En induisant une dysbiose, les antibiotiques peuvent entrainer des conséquences néfastes pour la santé. La principale complication à court terme est la modification du transit chez certains patients. Celle-ci se traduit le plus souvent par une diarrhée, le microbiote intestinal étant moins apte à remplir ses fonctions protectrices. Cette diarrhée associée aux antibiotiques est généralement d'intensité légère à modérée et son incidence varie selon l’âge, le type d’antibiotiques, le contexte… Elle peut toucher jusqu’à 35%9,10,11 des patients et chez les enfants, ce pourcentage peut atteindre jusqu'à 80%9. Dans 10 à 20% des cas, la diarrhée résulte d'une infection par Clostridioides difficile (C. difficile)11 : cette bactérie colonise le microbiote intestinal et va devenir pathogène sous l’influence de certains facteurs (une prise d’antibiotiques par exemple). Les conséquences cliniques sont variables, allant d’une diarrhée modérée à des symptômes beaucoup plus graves, voire la mort11.
35%
La diarrhée associée aux antibiotiques peut toucher jusqu'à 35 % des patients
80%
et jusqu'à 80% si les patients sont des enfants
4. Les antibiotiques peuvent engendrer des effets secondaires à plus long terme
La diarrhée associée aux antibiotiques n’est pas la seule manifestation de la dysbiose associée aux antibiotiques. Cette dernière serait responsable d’effets à plus long terme lorsqu’elle survient très tôt dans la vie. En effet, la période périnatale, qui se caractérise par le développement du microbiote intestinal et la maturation du système immunitaire, constitue une période particulièrement sensible12 : la dysbiose induite par la prise d’antibiotiques durant cette phase semble être un facteur de risque dans la survenue de maladies chroniques (obésité, diabète, asthme, maladies inflammatoires chroniques de l’intestin)13.
Image
5. L’usage inadapté des antibiotiques est responsable de la résistance aux antibiotiques
La résistance aux antibiotiques - ou antibiorésistance – correspond au fait qu’un traitement antibiotique ne soit plus efficace sur une infection bactérienne1. A quoi est-ce dû ? Les antibiotiques ne sont efficaces que sur les bactéries, et n’ont aucune action sur les virus (la grippe par exemple)14. Leur utilisation inadaptée (en cas d’infection virale par exemple) ou excessive - chez l’homme ou l’animal - accélère ce phénomène. La résistance aux antibiotiques entraine des hospitalisations plus longues, une hausse des dépenses de santé et une augmentation des décès. C’est pourquoi celle-ci constitue, à l’échelle mondiale, un enjeu majeur de santé publique1.
Image
6. Il existe, chaque année, une Semaine mondiale de sensibilisation à la résistance aux antimicrobiens
Chaque année, du 18 au 24 novembre, l’OMS organise la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens qui vise à mieux faire connaître le phénomène de résistance aux (sidenote:
Antimicrobiens
Catégories de médicaments regroupant les antibiotiques (action contre les bactéries), antiviraux (contre les virus), antiparasitaires (contre les parasites) et antifongiques (contre les champignons)
WHO Antimicrobial Resistance; Oct 2020) et d’encourager le grand public, les professionnels de santé et les responsables politiques à adopter de meilleures pratiques pour lutter contre l’émergence et la propagation des résistances. En tant qu’expert des microbiotes, le Biocodex Microbiota Institute se joint à cette initiative.
Si vous vous intéressez aux effets des antibiotiques sur votre santé et votre microbiote, ou si vous souhaitez en savoir plus sur la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens (WAAW), nous vous recommandons de consulter cette page dédiée :
La résistance aux antimicrobiens survient lorsque les bactéries, les virus, les parasites et les champignons évoluent et résistent aux effets des médicaments. En raison de cette pharmacorésistance, les antibiotiques et autres médicaments antimicrobiens perdent leur efficacité et les infections deviennent plus difficiles, voire impossibles à traiter, ce qui augmente le risque de propagation des maladies, de pathologie grave et de décès.
Organisée du 18 au 24 novembre, cette campagne invite le grand public, les professionnels de santé et les décideurs politiques à faire un usage raisonné des antibiotiques, des antiviraux, des antifongiques et des antiparasitaires afin de prévenir le développement accru de la résistance aux antimicrobiens.
A terme, elle pourrait menacer un siècle de progrès médical1. Véritable bombe sanitaire à retardement, la résistance antimicrobienne est dans le viseur de l’OMS qui organise chaque année, depuis 2015, la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens (18-24 novembre). L’Institut du Microbiote participe activement à cette initiative avec, pendant tout le mois de novembre, la diffusion et le partage de contenus exclusifs sur l’impact des antimicrobiens sur le microbiote intestinal. Tour d’horizon.
Côté pile, une extraordinaire découverte scientifique qui permet de sauver des millions de vies. Côté face, un usage excessif et parfois inadapté qui peut faire émerger de nombreuses résistances chez les microorganismes (que ce soit les bactéries, virus, parasites, champignons). Conséquence, les antimicrobiens, conçus pour guérir, sont de moins en moins efficaces et à terme, si aucune mesure n’est engagée, risque de ne plus parvenir à nous soigner contre les infections.
10 millions
La résistance aux antimicrobiens serait responsable de près de 10 millions de décès dans le monde d'ici 2050
Les résistances aux antimicrobiens seraient ainsi responsables, chaque année, de près de 700 000 décès dans le monde2. Si rien ne change, les maladies infectieuses pourraient devenir, en 2050, une des premières causes de mortalité dans le monde, en provoquant jusqu’à 10 millions de morts2.
Face à ce fléau, l’OMS organise la riposte mondiale. Depuis 2015, du 18 au 24 novembre, elle organise la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens qui s’évertue à mieux faire connaître ce phénomène mondial et à encourager le grand public, les professionnels de santé et les décideurs à adopter un usage raisonné des antibiotiques afin d’éviter que l’antibiorésistance ne gagne du terrain.
Véritable carrefour de connaissances dédié au microbiote, l’Institut du Microbiote est un partenaire actif de l’événement depuis 2020. Tout au long du mois de novembre, l’Institut vous invite, via des articles, actualités mais aussi vidéos d’experts et dossiers thématiques à approfondir vos connaissances et à découvrir les conséquences à moyen, long terme des antibiotiques sur le microbiote humain. Un exemple ? Malgré leur efficacité reconnue contre les bactéries (et inutiles en cas d’infection virale)3, ils entrainent souvent une dysbiose. Celle-ci est associé à certains troubles bien connus, tels que la diarrhée associée aux antibiotiques.
Fer de lance de l'arsenal thérapeutique moderne, les antibiotiques ont sauvé des millions de vie. En revanche, leur utilisation excessive et parfois injustifiée peut conduire à l'apparition de différentes formes de résistance chez les micro-organismes. Chaque année, l'Organisation Mondiale de la Santé (OMS) organise la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens (WAAW) afin de sensibiliser la population sur ce problème de santé publique. Lisez la page qui y est consacrée.
Mais ce n’est pas tout ! La prise d’antibiotiques est également soupçonnée d'augmenter le risque de plusieurs maladies chroniques4 (allergies, asthme, obésité, maladies inflammatoires chroniques de l’intestin…), et ceci particulièrement s’ils sont prescrits tôt dans l’enfance. Peut-on y remédier ? Oui ! En favorisant une prescription pertinente pour garantir le bon usage des antibiotiques ! Mais aussi en accompagnant le patient sur les risques de dysbiose associés à un usage excessif et inadapté des antibiotiques. Tous responsables, tous mobilisés pour réduire la résistance aux antimicrobiens !
Nous vous présentons le Professeur Sørensen, lauréat de la bourse internationale 2022 de la Biocodex Microbiota Foundation.
Son équipe a été la première à lancer une étude ambitieuse sur le résistome de 700 enfants, qui permettra de faire un pas de géant dans la compréhension de l'évolution et de la dissémination de la résistance aux antibiotiques dans l'intestin humain au cours de la petite enfance.
3 Improving Antibiotic Use. Material Developed by CDC Using CDC materials does not imply endorsement or recommendation by CDC, ATSDR, HHS or the United States Government
A terme, elle pourrait menacer un siècle de progrès médical.1 Véritable bombe sanitaire à retardement, la résistance antimicrobienne est dans le viseur de l’OMS qui organise chaque année, depuis 2015, la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens (18-24 novembre). L’Institut du Microbiote participe activement à cette initiative avec, pendant tout le mois de novembre, la diffusion et le partage de contenus exclusifs sur l’impact des antimicrobiens sur le microbiote intestinal. Tour d’horizon.
Côté pile, une extraordinaire découverte scientifique qui permet de sauver des millions de vies. Côté face, un usage excessif et parfois inadapté qui peut faire émerger de nombreuses résistances chez les microorganismes (que ce soit les bactéries, virus, parasites, champignons). Conséquence, les antimicrobiens, conçus pour guérir, sont de moins en moins efficaces et à terme, si aucune mesure n’est engagée, risque de ne plus parvenir à nous soigner contre les infections.
Image
Les résistances aux antimicrobiens seraient ainsi responsables, chaque année, de près de 700 000 décès dans le monde2. Si rien ne change, les maladies infectieuses pourraient devenir, en 2050, une des premières causes de mortalité dans le monde, en provoquant jusqu’à 10 millions de morts2.
10 millions
La résistance aux antimicrobiens serait responsable de près de 10 millions de décès dans le monde d'ici 2050
Face à ce fléau, l’OMS organise la riposte mondiale. Depuis 2015, du 18 au 24 novembre, elle organise la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens qui s’évertue à mieux faire connaître ce phénomène mondial et à encourager le grand public, les professionnels de santé et les décideurs à adopter un usage raisonné des antibiotiques afin d’éviter que l’antibiorésistance ne gagne du terrain.
Image
Véritable carrefour de connaissances dédié au microbiote, l’Institut du Microbiote est un partenaire actif de l’événement depuis 2020. Tout au long du mois de novembre, l’Institut vous invite, via des articles, actualités mais aussi vidéos d’experts, à découvrir les conséquences à moyen, long terme des antibiotiques sur le microbiote humain. Un exemple ? Prenez les antibiotiques. Malgré leur efficacité reconnue contre les bactéries (et inutiles en cas d’infection virale)3, ils perturbent l’équilibre au sein de notre microbiote intestinal. Ce déséquilibre, plus connu sous le nom de (sidenote:
Dysbiose
La « dysbiose » n’est pas un phénomène homogène : elle varie en fonction de l’état de santé de chaque individu. Elle est généralement définie comme une altération de la composition et du fonctionnement du microbiote, provoquée par un ensemble de facteurs environnementaux et liés à l’individu, qui perturbent l’écosystème microbien.
Levy M, Kolodziejczyk AA, Thaiss CA, et al. Dysbiosis and the immune system. Nat Rev Immunol. 2017;17(4):219-232.), est associé à certains troubles bien connus, tels que la diarrhée associée aux antibiotiques. Mais ce n’est pas tout ! La prise d’antibiotiques est également soupçonnée d'augmenter le risque de plusieurs maladies chroniques (allergies, asthme, obésité, maladies inflammatoires chroniques de l’intestin…), et ceci particulièrement s’ils sont prescrits tôt dans l’enfance.
Image
Peut-on y remédier ? Oui ! D’abord en privilégiant un usage juste et approprié. Ne prenez pas ces médicaments sans prescription d’un professionnel de santé. Respectez la dose, la posologie, la durée de votre traitement et ne le partagez pas avec une autre personne.4
Si vous vous intéressez aux effets des antibiotiques sur votre santé et votre microbiote, ou si vous souhaitez en savoir plus sur la Semaine mondiale de sensibilisation à la résistance aux antimicrobiens (WAAW), nous vous recommandons de consulter cette page dédiée:
La résistance aux antimicrobiens survient lorsque les bactéries, les virus, les parasites et les champignons évoluent et résistent aux effets des médicaments. En raison de cette pharmacorésistance, les antibiotiques et autres médicaments antimicrobiens perdent leur efficacité et les infections deviennent plus difficiles, voire impossibles à traiter, ce qui augmente le risque de propagation des maladies, de pathologie grave et de décès.
Organisée du 18 au 24 novembre, cette campagne invite le grand public, les professionnels de santé et les décideurs politiques à faire un usage raisonné des antibiotiques, des antiviraux, des antifongiques et des antiparasitaires afin de prévenir le développement accru de la résistance aux antimicrobiens.
Revue de presse Par le Pr. Markku Voutilainen Faculté de médecine de l'Université de Turku ; gastro-entérologie, Hôpital universitaire de Turku, Finlande
Une étude récente montre que l’huile de noix de coco aide au maintien d’un cuir chevelu sain en agissant favorablement sur son microbiote. Les chercheurs ont comparé l’impact d’une application d’huile de coco versus un shampoing neutre sur le microbiote bactérien et fongique du cuir chevelu de 140 femmes avec ou sans pellicules. Dans le cuir chevelu des femmes avec des pellicules, on retrouvait des populations beaucoup plus importantes de champignons Malassezia, qui accélèreraient le développement des pellicules et de l’inflammation. A contrario, une autre espèce de champignons, M. globosa, colonisait en abondance le cuir chevelu des femmes qui ne présentaient ni pellicules ni démangeaisons. Le traitement à l’huile de noix de coco a permis d’augmenter la proportion de M. globosa par rapport aux autres groupes de Malassezia, vers un ratio similaire aux cuirs chevelus en bonne santé. Bien qu’aucune différence significative n’ait été observée entre le microbiote bactérien du groupe sain et celui présentant des pellicules, le traitement à l’huile de coco a permis, dans les deux groupes, une augmentation des bactéries impliquées dans le métabolisme de la biotine. Cette vitamine B est essentielle au maintien d’une peau et d’un cuir chevelu en bonne santé. Elle est également connue pour réduire l’inflammation. Des études complémentaires s’avèrent nécessaires pour comprendre les mécanismes sous-jacents mais, pour les chercheurs, l’effet positif de l’huile de coco sur la composition et la fonction des communautés microbiennes serait la première étape vers une restauration à plus long terme d’un cuir chevelu sain.
Revue de presse Par le Pr. Markku Voutilainen Faculté de médecine de l'Université de Turku ; gastro-entérologie, Hôpital universitaire de Turku, Finlande
Une étude montre que grâce à des endolysines recombinantes du type 1,4-bêta- N-acétylmuramidase, provenant de prophages au sein du génome de Gardnerella, il est possible d’éliminer le biofilm bactérien responsable de la vaginose bactérienne, sans endommager les bactéries bénéfiques du microbiote vaginal. Pour cela, les auteurs ont créé plusieurs endolysines - des enzymes des bactériophages, qui lysent la paroi des bactéries - modifiées par réarrangement de domaine. Ils ont testé leur activité bactéricide sur des souches de Gardnerella par rapport aux endolysines sauvages. Les endolysines recombinantes avaient une activité bactéricide 10 fois plus élevée que celles de type sauvage. La plus active d’entre elles, baptisée PM-477, a été testée contre un panel de 20 souches de Gardnerella au sein de 4 espèces (G. vaginalis, G. leopoldii, G. piotii et G. swidsinski) et a montré une efficacité supérieure par rapport aux antibiotiques testés (métronidazole, tinidazole, clindamycine). En revanche, PM-477 n’a eu aucun effet sur les lactobacilles bénéfiques ou d’autres espèces de bactéries vaginales. Les auteurs en ont déduit que l’endolysine PM-477 était hautement sélective envers Gardnerella et tuait les souches de chacune des quatre principales espèces, sans toucher les lactobacilles bénéfiques ou autres espèces typiques du microbiote vaginal. Ils ont confirmé cette observation par microscopie, dans des co-cultures de Gardnerella et lactobacilles. Pour aller plus loin et analyser l’efficacité de PM-477 dans un environnement physiologique proche de la situation in vivo, les chercheurs ont traité les prélèvements vaginaux de 15 patientes atteintes de vaginose bactérienne et les ont analysés par hybridation in situ en fluorescence (FISH). Ils ont montré que dans 13 cas sur 15, PM-477 tuait la bactérie Gardnerella et dissolvait physiquement les biofilms sans affecter le microbiote vaginal. Pour les auteurs, l’utilisation d’endolysines serait une option thérapeutique prometteuse pour lutter contre la vaginose bactérienne et se passer des antibiotiques.