What is putting me in such a good mood today?
Considered the “second brain”, our gut is constantly talking to our brain and vice versa. This is known as the gut-brain axis. The malfunctioning of this axis could be involved in some neuropsychiatric disorders: anxiety, depression and attention disorders... This is still under review: modulating the gut microbiota through diet or the administration of probiotics could prevent, even treat these conditions.
In addition, some foodstuffs are even supposedly able to trigger a “feel-good effect”.
So what is the link between your gut flora and your mood? Answer below!
The gut microbiota Mood disorders Anxiety disorders Have you heard of "dysbiosis"? Diet
The dialogue between our brain and our gut regulates our mood
With its 200 million neurons and its billions of gut bacteria, our gut deserves its nickname “second brain”. It is highly active, and contributes to both our physical and mental health.1
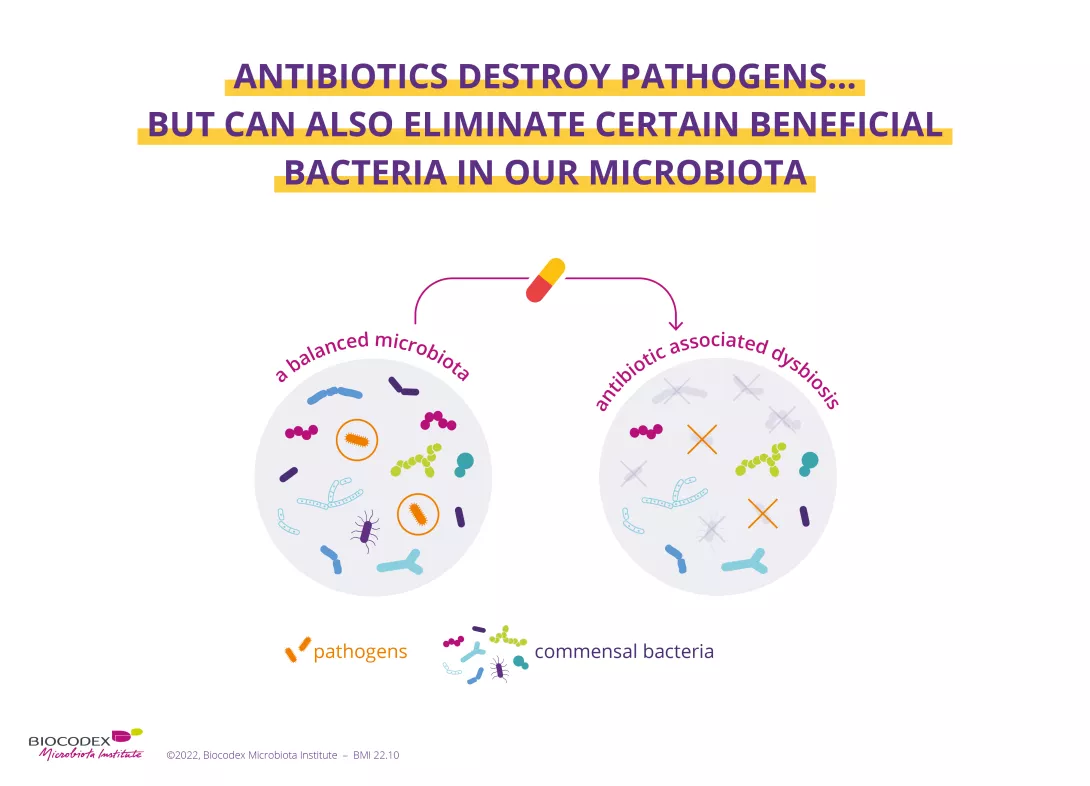
Our digestive tract and brain are constantly talking to each other, but communication between the two may be disrupted when our gut microbiota is degraded and an inflammatory process is set up.
200 millions neurons on the gut
1 billion of gut bacteria
The disruption of our gut microbiota, also known as dysbiosis, is allegedly involved in various mental health disorders (depression, anxiety).2 In fact, some studies conducted in humans (albeit only a few) appear to have demonstrated a significant reduction in the richness of the gut flora in patients with mental disorders.3

Gut-brain axis: how does it work?
The gut bacteria talk to our brain by producing molecules called “neurotransmitters”; the most well-known, serotonin and dopamine, regulate our mood.4 According to scientists, these (sidenote: Neurotransmitters Specific molecules that enable communication between the neurons (the nerve cells in the brain), as well as with the bacteria in the microbiota. They are produced by the individual’s cells and by the bacteria in the microbiota. Baj A, Moro E, Bistoletti M, Orlandi V, Crema F, Giaroni C. Glutamatergic Signaling Along The Microbiota-Gut-Brain Axis. Int J Mol Sci. 2019;20(6):1482. ) act on the cells in the gut wall. In this way the message reaches the brain via the neurons of the digestive tract.5

Another facilitator of information: (sidenote: Short chain fatty acids (SGFA) Short chain fatty acids are a source of energy (fuel) for an individual’s cells. They interact with the immune system and are implicated in communication between the gut and the brain. Silva YP, Bernardi A, Frozza RL. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front Endocrinol (Lausanne). 2020;11:25. ) (SCFA). These biological substances, some of which are good for our bodies, are produced by the bacteria in our colon during fermentation of dietary fibers. They also play a key role in the connections between the two organs by acting directly on the brain.6
Gut microbiota: still many things to be discovered
A good mood and dark chocolate: an 85% happy marriage!
Did you know? Some foods, such as chocolate, can regulate our mood.
A study7 recently showed that the polyphenols found in large quantities in cocoa may have a positive action on the gut flora slowing down the growth of (sidenote: Pathogen A pathogen is a microorganism that causes, or may cause, disease. Pirofski LA, Casadevall A. Q and A: What is a pathogen? A question that begs the point. BMC Biol. 2012 Jan 31;10:6. ) bacteria and encouraging the development of beneficial ones.

It is in particular the consumption of dark chocolate with 85% cocoa solids that supposedly increases the microbial diversity in our gut and sets up a virtuous circle with the brain, resulting in a long-term positive effect on our mood.
Chocolate-lovers, you can finally stop feeling guilty!