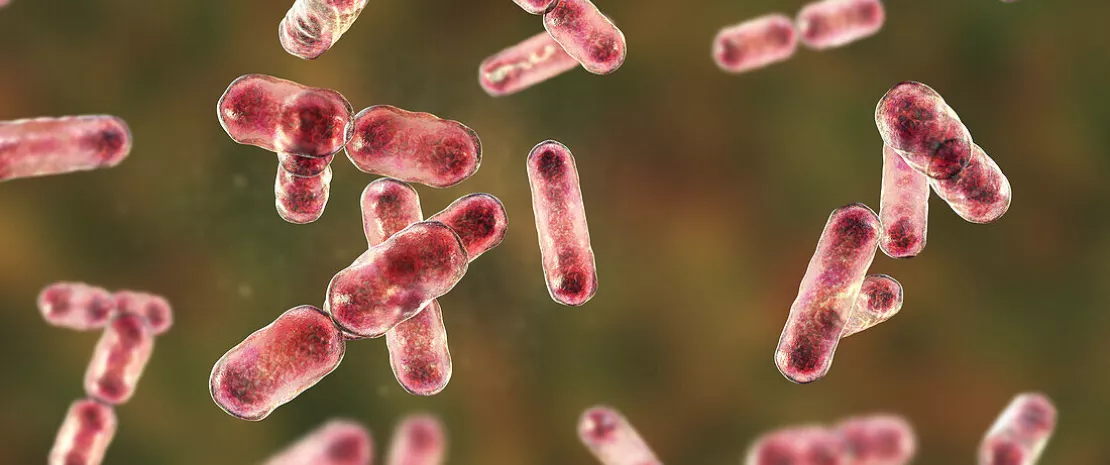
Moins de diversité, moins de micro-organismes bénéfiques et davantage de pathogènes opportunistes : on sait que le microbiote intestinal évolue avec l’avancée en âge. Mais qu’en est-il chez les centenaires, qui ont réussi à avoir une vie plus longue et à échapper à diverses maladies chroniques et infections ? Pour étudier la relation entre microbiote intestinal et longévité, des chercheurs d’institut de recherche chinois ont comparé le microbiote intestinal de 1 575 personnes âgées de 20 à 117 ans, vivant dans la même province de Guangxi en Chine : 314 jeunes adultes (20-44 ans), 277 adultes (45-65 ans), 386 seniors (66-85 ans), 301 nonagénaires (90-99 ans) et 297 centenaires (100 à 117 ans). Les résultats de cette nouvelle étude sur le vieillissement du microbiote ont été publiés dans Nature Aging.
100 ans et un microbiote de 20 ans
Bilan des courses : la diversité des espèces du microbiote intestinal recule avec les années, pour être au plus bas chez les seniors de 66-85 ans. Mais de manière surprenante, elle réaugmente chez les nonagénaires et les centenaires : ces derniers présentent les flores les plus riches, au même titre que les individus les plus jeunes. Néanmoins, les chercheurs estiment que ce n’est pas tant la diversité d’espèces que l’homogénéité des abondances relatives des diverses espèces qui pourrait être liée à la longévité.
Autres signes de jeunesse et de bonne santé du microbiote des seniors de 100 ans : d’une part, une présence accrue des Bacteroïdetes potentiellement bénéfiques, comparativement aux seniors et nonagénaires ; d’autre part, une présence réduite en bactéries potentiellement pathogènes, notamment inflammatoires.
Une singularité qui se renforce après 100 ans
Ces premiers résultats suggérant qu'une signature spécifique pourrait caractériser le microbiote intestinal des personnes âgées de plus de 100 ans, les chercheurs ont également mené une étude longitudinale des altérations microbiennes intestinales chez 45 des 297 centenaires, chez lesquels un second échantillon de selle a été collecté en moyenne 1,5 an plus tard. Le microbiote intestinal des seniors de 100 ans gagne en uniformité des abondances relatives des espèces présentes, avec en outre une baisse de la variation interindividuelle et une stabilité de la population des Bacteroïdetes. L’uniformité des abondances au début de l’étude s’avère d’ailleurs corrélée à la stabilité du microbiote intestinal des centenaires sur les 1,5 années, laissant penser que cet équilibre entre espèces pourrait protéger la flore intestinale d’un dérèglement et de son vieillissement.
Selon les chercheurs, les seniors de 100 ans se distingueraient par des profils de microbiotes spécifiques, jouissant d’un équilibre entre espèces non seulement élevé pour leur âge mais qui continue aussi à croître, et d’une abondance stable de Bacteroïdetes. Malgré les années, leur flore intestinale conserve une grande similarité avec celle de jeunes adultes.





 Dépression : un diagnostic plus précis grâce au microbiote intestinal ?
Dépression : un diagnostic plus précis grâce au microbiote intestinal ?
 Dépression : vers une confirmation du dialogue intestin-cerveau ?
Dépression : vers une confirmation du dialogue intestin-cerveau ?