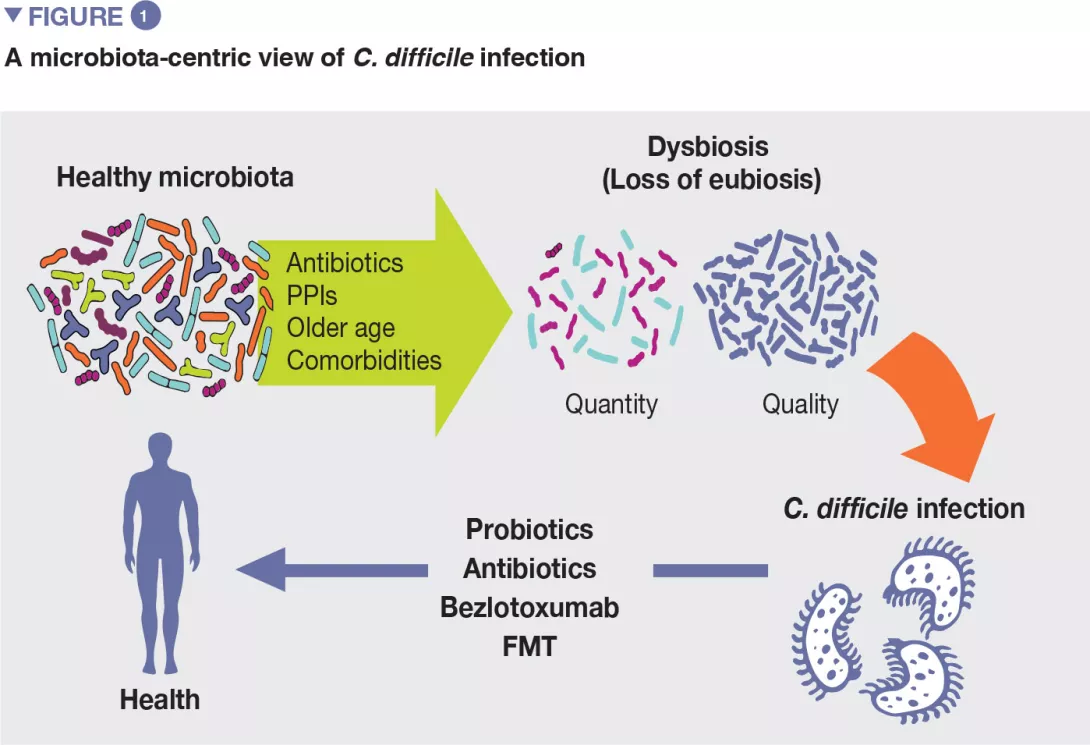
If antibiotics remain today essential molecules in the therapeutic arsenal, it is also necessary to take into account their undesirable effects on the gut microbiota, as a considerable body of evidence supports the association between their use and many dysbiosis-associated diseases, including CDI.[5]
First, antibiotics may kill commensal bacteria that may have a direct action against C. difficile (by secreting a number of bacteriocins) and also compete with the pathogen for nutrients (e.g., sialic acid and succinate). Moreover, there is also an indirect protective role of commensal bacteria through the regulation of bile acids.
Recently, Clostridium scindens was associated with resistance to C. difficile colonisation. It has a bile acid inducible operon which is able to encode dehydroxylating enzymes that convert primary bile acids into secondary bile acids. Primary bile acids promote the germination of C. difficile spores, while secondary bile acids are able to inhibit this process.[6]
The use of systemic antibiotics is the most relevant modifiable risk factor for the development of CDI. Healthy gut microbiota can determine the successful colonisation of the large bowel by C. difficile or not, by direct and indirect pathways. In principle, the imbalance of healthy gut microbiota by broad-spectrum antibiotics may bring several consequences that drive to CDI.
As a corollary of this evidence, patients with recurrent CDI are known to have an imbalanced microbial profile, with higher relative abundance of detrimental bacterial families as Enterobacteriaceae and Veillonellaceae and lower relative abundance of beneficial families, including Ruminococcaceae, Bacteroidaceae and Lachnospiraceae.
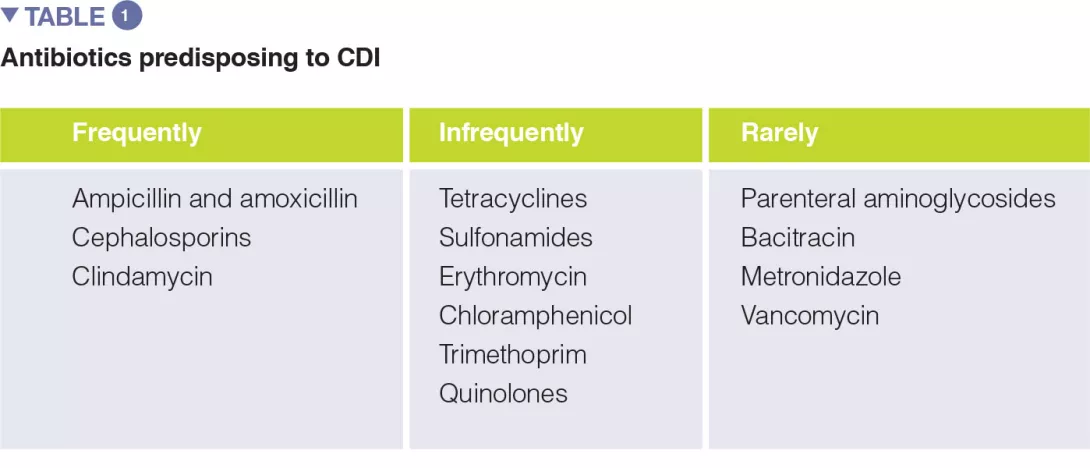
A number of systematic reviews, alone or with meta-analysis, have assessed the relevance of different antibiotic classes in CDI development. In the earliest meta-analysis (1998), antibiotics use was associated with a 6-fold increase in the risk of developing CDI, and the highest risk was observed for fluoroquinolones, clindamycin, cephalosporins. Moreover, the use of antibiotics was found to be an independent predictor of CDI recurrence (relative risk 1.76). One of the key factors to prevent CDI is represented by the antibiotic stewardship approach, so the knowledge of the CDI risk for different antibiotic classes is of paramount importance (Table 1).
The use of the following antibiotics is associated with a 2-fold higher risk of CDI among inpatients: clindamycin, cephalosporins, carbapenems, fluoroquinolones, trimethoprim, sulphonamides. In the community setting, respectively, antibiotics were found to have different risk levels for CDI development or recurrence, including: clindamycin (risk increased of 8 to 20 times), cephalosporins and fluoroquinolones (3-5 times increase), macrolides (2-3 times increase).[5]