Un nuevo estudio revela que el trasplante de microbiota fecal mejora los síntomas del síndrome de colon irritable y la calidad de vida de los pacientes, incluso un año después del tratamiento, pero que es imprescindible que el donante goce de buena salud.
Dolores, calambres, gases, diarrea, estreñimiento... El síndrome de colón irritable es una dolencia que se manifiesta por un conjunto de síntomas abdominales que van y vienen a lo largo de la vida del paciente. Estos síntomas pueden empeorar con el estrés, los cambios emocionales o algunos alimentos... y alteran considerablemente la calidad de vida del paciente. Si no existe ningún problema anatómico o estructural del intestino de las personas con este síndrome, la microbiota intestinal, en cambio, suele tener algo que ver.
¿Un solo donante con una supermicrobiota?
Los investigadores de este estudio han probado en un ensayo clínico la eficacia de un trasplante de microbiota fecal en el que utilizaron las muestras de heces de un único hombre caucásico de 36 años1 que cumplía todos los requisitos para ser un «superdonante»: gozar de buena salud, IMC normal, realizar ejercicio de forma regular, haber nacido por vía vaginal y haber sido amamantado. Además, no tomaba ninguna medicación y solo había sido tratado con antibióticos tres veces en su vida e ingería regularmente complementos alimentarios. En este ensayo clínico, el trasplante de la microbiota fecal ha demostrado ser eficaz en pacientes con síndrome de colon irritable. Sin embargo, estos resultados solo se observaron 3 meses después del trasplante y quedan muchas preguntas por resolver, la más importante saber si el efecto clínico del trasplante se mantiene a largo plazo. Los investigadores han realizado un seguimiento a los pacientes del estudio durante un año tras el estudio.
Tras 1 año, los beneficios siguen presentes
La mayoría de los pacientes que había respondido bien al trasplante de microbiota fecal 3 meses después mantenían esa respuesta 1 año después. Otro resultado motivador es la mejora de los síntomas abdominales, la fatiga y la calidad de vida tres meses después del trasplante. Lo que es aún mejor, entre el 32 y el 45 % de los pacientes, según el grupo, han experimentado una remisión total de los síntomas durante el año siguiente. El análisis completo de la microbiota intestinal de los pacientes ha mostrado cambios en el perfil bacteriano intestinal y una reducción significativa del índice de disbiosis.
En conclusión, el trasplante de microbiota fecal de un «superdonante» permitiría restablecer la microbiota intestinal y disminuir los síntomas en pacientes que padecen síndrome de colon irritable.
La disbiosis consiste en la ruptura del delicado equilibrio entre los miles de millones de microorganismos que conforman la microbiota humana y su relación con nuestro cuerpo. Genética, alimentación poco equilibrada, antibióticos... Las causas de la disbiosis son variadas y, en su mayoría, múltiples. Actualmente, la investigación científica ha demostrado que la disbiosis de la microbiota intestinal, la más estudiada, así como también de otras microbiotas, como la vaginal, la cutánea o la pulmonar, está relacionada con diferentes enfermedades, desde el síndrome del colon irritable hasta trastornos metabólicos, como la obesidad, pasando incluso por la sinusitis crónica y el eccema. ¿Cómo puede desequilibrarse la microbiota? ¿Cuáles son las consecuencias de la disbiosis para nuestra salud? ¿Cómo podemos recuperar el equilibrio de la microbiota?
En primer lugar, volvamos al propio término: «disbiosis». La etimología de este término científico es muy sencilla. En griego, la palabra bios significa «vivo» y el prefijo dis- significa «malo».
La «disbiosis» puede definirse como una alteración en la composición y el funcionamiento de la microbiota. Dicha alteración viene derivada de una combinación de factores ambientales y específicos de cada persona1.
Puesto que los microorganismos colonizan todo nuestro cuerpo, la disbiosis puede observarse en múltiples lugares:
En la microbiota intestinal: se han asociado varias enfermedades a la disbiosis intestinal, como la diarrea postantibiótica14, la gastroenteritis17, el cólico del lactante44…
En la microbiota cutánea: la disbiosis a menudo se asocia a procesos patológicos (acné45, dermatitis atópica46).
En la microbiota vaginal: una disbiosis vaginal se asocia a una vaginosis bacteriana1, una candidiasis47, una menor fertilidad48 o un mayor riesgo de parto prematuro1.
En la microbiota ORL (oreja, nariz, garganta): varias enfermedades pueden asociarse a un desequilibrio de la microbiota oral, auricular o nasofaríngea.
En la microbiota pulmonar: la disbiosis puede intervenir en la aparición de infecciones respiratorias invernales49, asma50 o fibrosis quística51.
En la microbiota urinaria: se ha demostrado en estudios publicados hasta la fecha el papel que desempeña la microbiota urinaria en las infecciones urinarias52.
La disbiosis sobre la mesa: la microbiota intestinal
La microbiota intestinal es la principal microbiota del cuerpo humano2. Acoge al menos a 1000 especies diferentes3 de microorganismos, como bacterias, hongos y virus. El filo Firmicutes (que incluye las famosas «bacterias buenas» lactobacilos) y el filo Bacteroidetes combinados suponen entre el 70 y el 90 % de la comunidad bacteriana de nuestro intestino2-4. Nuestra microbiota también contiene actinobacterias, como bifidobacterias, conocidas por sus efectos beneficiosos. Otros microorganismos de nuestra microbiota pueden hacer que enfermemos. Estos se conocen como «posibles (sidenote:
Patógeno
un patógeno es un microorganismo que causa, o puede causar, una enfermedad
Pirofski LA, Casadevall A. Q and A: What is a pathogen? A question that begs the point. BMC Biol. 2012 Jan 31;10:6.)», pero son una minoría2. La disbiosis da como resultado uno o más de los siguientes fenómenos:
Las proporciones entre estas grandes familias de bacterias cambian significativamente, en particular con una pérdida de lactobacilos y bifidobacterias5.
Los microorganismos útiles que viven habitualmente en nuestra microbiota (llamados «comensales») se reducen o desaparecen1.
La diversidad de microorganismos presentes en la microbiota se agota: hay menos especies diferentes5;
Los microorganismos potencialmente patógenos de la microbiota se multiplican1,5.
Consecuencias: nuestra microbiota se vuelve débil y las bacterias «malas» se apoderan de las «buenas» 2. Protege nuestro cuerpo con menos facilidad contra las agresiones y cumple con menor eficacia sus funciones esenciales para mantener nuestra forma y nuestra salud.1,6.
¿Cuáles son las características de la disbiosis?
Aunque la disbiosis en sí misma no se considera una enfermedad, se ha relacionado con varios problemas de salud y puede contribuir a la aparición o el empeoramiento de ciertas afecciones.
1000
Acoge al menos a 1000 especies diferentes de microorganismos.
Desequilibrio de la microbiota específica de cada persona
Sin embargo, ¡la disbiosis no es un término universal que se pueda aplicar a todas las personas y en todas las circunstancias!1 De hecho, se ve afectada por nuestra genética y microorganismos (definición: organismos vivos que son demasiado pequeños para ser vistos a simple vista. Incluyen bacterias, virus, hongos, arqueas, protozoos, etc... Estos microorganismos, que se denominan comúnmente « (sidenote: https://microbiologysociety.org/why-microbiology-matters/what-is-microbiology.html)»), colonizaron nuestro cuerpos durante los primeros años de vida. Asimismo, la composición de nuestra microbiota es personal. Varía tanto entre individuos que podría ser tan exclusiva como una huella digital.7 Pero también puede cambiar dependiendo de nuestra edad, nuestro estado de salud, la aparición del estrés, nuestra alimentación, el lugar donde vivimos, los medicamentos que tomamos…8 Por lo tanto, cada uno de nosotros puede tener «su» disbiosis cuando nuestra microbiota se desequilibra y no funciona correctamente en nuestro organismo.1
Entonces, ¿qué significa tener una microbiota equilibrada?
El prefijo dis de disbiosis se opone a eu («bueno») o sim («con»). Por tanto, hablamos de «eubiosis» o «simbiosis» cuando nuestra microbiota está en buen estado : interactúa en armonía con nuestro organismo y su comunidad microbiana está equilibrada.1
De hecho, existe un beneficio mutuo entre los miles de microorganismos que habitan nuestra microbiota y nuestro cuerpo.9 Todos salen ganando: el cuerpo proporciona «alimento y refugio» a los microorganismos de la microbiota. A cambio, estos participan en muchas funciones importantes de nuestro cuerpo, como la digestión, la asimilación de nutrientes, la protección de la impermeabilidad de la pared intestinal y la lucha contra gérmenes no deseados.2,8,10 ¡Es todo un trabajo en equipo!
Los diferentes microorganismos de la comunidad de nuestra microbiota, incluidos aquellos que pueden ser patógenos, están presente en número y proporciones adecuadas para convivir de manera pacífica y garantizar sus funciones beneficiosas para el organismo. Sin embargo, este sutil equilibrio entre los ecosistemas microbianos de nuestro cuerpo puede romperse: la eubiosis se transforma en disbiosis.8
¿Qué causa una disbiosis?
Tal como indica su definición, la disbiosis aparece por el efecto de varios factores muy diferentes, a menudo entrelazados.5. No obstante, podemos distinguir entre:
Aquellos que están relacionados con el propio individuo, tales como:
La genética.1
La edad.11
Ciertas afecciones y lesiones. 1
Aquellos que están relacionados el entorno, tales como:
La ingesta de medicamentos: antibióticos, antiinflamatorios...2,5
Las infecciones.
El estilo de vida: alimentación desequilibrada o cambios de alimentación, estrés, tabaquismo, una higiene inadecuada...1, 5,8 .
Los antibióticos: irremplazables, pero alteran la microbiota
Los antibióticos han supuesto uno de los progresos terapéuticos más importantes del siglo XX. Tras el descubrimiento de la penicilina en 1928, se han salvado millones de vidas.13 Sin embargo, al destruir los gérmenes dañinos y también las bacterias «buenas», se desequilibra la microbiota. A corto plazo, la disbiosis generada por los antibióticos puede derivarse en diarrea14 o micosis vaginal 15. También se sospecha que la disbiosis intestinal inducida por antibióticos tiene un impacto a largo plazo, especialmente cuando se toman antibióticos durante la infancia, al aumentar el riesgo de diversas enfermedades crónicas como la obesidad o las alergias 16 ...
Infecciones: cuando atacan los microbios
Durante infecciones como la gastroenteritis vírica o la intoxicación alimentaria por salmonela, los microbios dañinos y agresivos invaden la microbiota humana. No provienen de la microbiota, sino del exterior, y se transmiten, por ejemplo, por las manos o alimentos contaminados. Estas infecciones provocan una fuerte reacción de nuestro sistema inmune, una inflamación del intestino y diarrea. Todo ello provoca alteraciones bruscas en el equilibrio de nuestra flora intestinal. Además, los microbios implicados en estas infecciones también pueden promover el desarrollo de otras bacterias potencialmente patógenas ya presentes en la microbiota. Por lo tanto, las infecciones causan disbiosis, ¡de la que se benefician todas las bacterias dañinas !1,12,17,18
Alimentación: el equilibrio de la microbiota en nuestros platos
Lo que comemos afecta a la microbiota a lo largo de nuestra vida. Un cambio repentino en la dieta, en composición o cantidad, puede desencadenar disbiosis. Pero no solo eso: si las variaciones habituales de nuestros menús de un día para otro solo provocan modificaciones transitorias de la microbiota, nuestro tipo de alimentación puede modificar el ecosistema digestivo de manera permanente5 y puede, a largo plazo, ser un factor de disbiosis. Por tanto, los estudios sugieren que una dieta «al estilo occidental» rica en grasas, azúcares y proteínas puede provocar desequilibrios en la microbiota intestinal, mientras que una dieta variada rica en frutas y verduras podría protegerla.1,19
Antibióticos
Han salvado millones de vidas, pero su uso excesivo e inapropiado supone actualmente un grave peligro para la salud, sobre todo debido a la resistencia a los antibióticos y a la disbiosis de la microbiota. Todos los años, la OMS organiza la Semana mundial de concienciación sobre la RAM (WAAW) para aumentar la sensibilización sobre la resistencia antimicrobiana. Echemos un vistazo a esta página dedicada al tema.
¿Qué es la Semana mundial de concienciación sobre la RAM?
Cada año, desde 2015, la OMS organiza la Semana mundial de concienciación sobre la RAM (WAAW por sus siglas en inglés), cuyo objetivo es fomentar la sensibilización sobre la resistencia global antimicrobiana.
La resistencia antimicrobiana se produce cuando las bacterias, los virus, los parásitos y los hongos cambian con el tiempo y dejan de responder a los medicamentos. Debido a la resistencia a los fármacos, los antibióticos y otros medicamentos dejan de ser efectivos y las infecciones son cada vez más difíciles o incluso imposibles de tratar, lo que aumenta el riesgo de propagación de las enfermedades, de sufrir dolencias graves y de muerte.
Esta campaña, que se desarrolla del 18 al 24 de noviembre, anima al público en general, a los profesionales de la salud y a los responsables políticos a usar los antibióticos, los antivirales, los antifúngicos y los antiparasitarios con cuidado, para prevenir la aparición de nuevas resistencias antimicrobianas.
Disbiosis: ¿causa o consecuencia de una enfermedad?
Se ha demostrado en numerosos estudios que comparaban la microbiota de personas sanas con la de personas enfermas que la disbiosis está asociada a diversas afecciones crónicas: enfermedades intestinales, como el síndrome del colon irritable o la enfermedad de Crohn, así como obesidad, alergias, asma y determinados cánceres1. Pero ¿es la disbiosis la que provoca la enfermedad o es la enfermedad la que causa la disbiosis? Para los científicos, la respuesta no siempre es obvia, pero actualmente es objeto de mucha investigación.
Con el fin de arrojar luz, los investigadores lanzaron Homo symbiosus en 2019, un gran proyecto de investigación que tiene como objetivo determinar mejor por qué y cómo tantas enfermedades crónicas están relacionadas con la disbiosis intestinal. Los investigadores plantearon la hipótesis de que todos «estos fenómenos de disbiosis intestinal, proliferación de microbios, inflamación y debilitamiento de la pared intestinal» se apoyan entre sí10.
La disbiosis de la microbiota intestinal se asocia a diversas enfermedades: no solo dolencias gastrointestinales, sino también metabólicas22, alérgicas23 e incluso mentales24. No obstante, el cuerpo humano también alberga ecosistemas microbianos específicos en la piel25, las vías urinarias26, la vagina27 o la boca28 y los pulmones29, cuya composición puede desequilibrarse y relacionarse con determinadas enfermedades.
¿Cómo podemos recuperar el equilibrio de la microbiota?
Normalmente, después de un episodio de disbiosis, la microbiota es capaz de recuperar de forma natural su equilibrio inicial (aunque nunca recuperamos por completo la composición inicial): se dice que es «resiliente»30. Pero a veces esta «rebiosis», es decir, la vuelta al equilibrio microbiano, puede llevar tiempo: por ejemplo, incluso en un adulto sano, puede tardar hasta seis meses después de tomar un antibiótico31. Por último, la disbiosis a veces provoca un estado de desequilibrio que perdurará y se mantendrá en el tiempo sin poder recuperarse por completo, lo que puede ser perjudicial para la salud1.
¿Qué hacemos ante una disbiosis? Disponemos de varias soluciones para restablecer el equilibrio de la microbiota y recuperar una mejor salud.
Los probióticos: microorganismos de refuerzo beneficiosos
Los probióticos son «microorganismos vivos que, cuando se administran en la cantidad correcta, resultan beneficiosos para la salud del que los toma».30,31Aquí encontrará una página sobre los probióticos, cómo funcionan, cómo se elaboran, cómo elegirlos bien... Consulte nuestra página sobre los probióticos.
Los prebióticos: su función, alimentar nuestra microbiota
Derivados principalmente de fibras alimentarias (fructooligosacáridos, galactooligosacáridos, inulina, etc.), los prebióticos son nutrientes o sustratos no digeribles, utilizados por los microorganismos de la microbiota y que tienen efectos favorables en la salud.32,33Aquí encontrará más información sobre su forma de actuar sobre la microbiota . Los productos específicos que combinan probióticos y prebióticos se denominan simbióticos.34,35
Una alimentación saludable para preservar su buen funcionamiento
Lo que comemos, tanto la calidad como la diversidad de nuestra alimentación, contribuye al equilibrio de nuestra microbiota intestinal38,39, pero también influirá en su composición y, a su vez, será responsable de provocar determinadas enfermedades22. ¡No dude en consultar a su médico de cabecera o nutricionista para obtener más información sobre los alimentos que tienen efectos beneficiosos o nocivos para mantener su intestino sano y en buena forma 40!
El trasplante de microbiota, una técnica esperanzadora
Al igual que otros órganos, la microbiota se puede trasplantar de un individuo a otro para intentar restablecer el equilibrio de su ecosistema microbiano41,42. Por el momento, este enfoque terapéutico, conocido como trasplante de microbiota fecal (TMF), está bien documentado para la microbiota intestinal; sin embargo, solo está autorizado para el tratamiento de infecciones recurrentes por Clostridioides difficile41. Es objeto de una intensa investigación para otras patologías intestinales41. Para la vagina, el trasplante de microbiota vaginal (TMV) se está sometiendo a ensayos actualmente y podría ser un tratamiento prometedor para la vaginosis bacteriana recurrente o resistente43. Los estudios sobre el trasplante de microbiota cutánea son aún poco frecuentes, pero los resultados iniciales son alentadores44,45.
Un formidable avance médico víctima de su éxito. Desde su descubrimiento a principios del siglo xx, los antibióticos salvaron millones de vidas. Pero el uso masivo y a veces inadecuado de estos tratamientos los hace cada vez menos eficaces para tratar las infecciones. ¡El resultado es que, en la actualidad, numerosas bacterias son resistentes! Pero el empleo excesivo o inadecuado no es el único responsable de la resistencia a los antibióticos.
Un estudio1revela quelos viajes internacionales favorecen la adquisición de genes de resistencia a los antibióticos y podrían contribuir a la propagación de la resistencia a estos medicamentos. Embarque inmediato para algunas explicaciones.
Como cada año desde 2015, la Semana mundial de concienciación sobre la RAM (WAAW) nos sensibiliza sobre el aumento de la resistencia a los (sidenote:
Antibiorresistencia
Se habla de resistencia a los antimicrobianos cuando las bacterias, los virus, los hongos y los parásitos ya no responden a los medicamentos debido a su evolución en el tiempo. Los antibióticos y otros medicamentos antimicrobianos pierden su eficacia y las infecciones se vuelven cada vez más difíciles e incluso imposibles de tratar. Esta resistencia a los microbios aumenta el riesgo de propagación, de padecer una forma grave de la enfermedad e incluso de fallecimiento. La antibiorresistencia es la resistencia de las bacterias a los antibióticos.
Fuente: Résistance aux antimicrobiens. 26 Octobre 2020. ) (también llamada antibiorresistencia). Este fenómeno, debido al mal uso o a un consumo excesivo de antibióticos, designa la capacidad de una bacteria a resistirse a la acción de un antibiótico. En 2020, la WAAW amplió el alcance de su campaña a los (sidenote:
Antimicrobianos
Medicamentos —como los antibióticos, los antivirales, los antifúngicos y los antiparasitarios— utilizados para prevenir y tratar las infecciones en los seres humanos, los animales y los vegetales.
WHO Antimicrobial Resistance; Nov 2021): antivirales, antifúngicos, antiparasitarios…, es decir, a los medicamentos indispensables para luchar contra los (sidenote:
Microorganismos
Organismos vivos que son demasiado pequeños para verlos a simple vista. Incluyen las bacterias, los virus, los hongos, las arqueas, los protozoos, etc. y se llaman comúnmente «microbios».
Fuente: What is microbiology? Microbiology Society. ) (sidenote:
Patógeno
un patógeno es un microorganismo que causa, o puede causar, una enfermedad
Pirofski LA, Casadevall A. Q and A: What is a pathogen? A question that begs the point. BMC Biol. 2012 Jan 31;10:6.) - Al reducir las posibilidades de tratamiento de las infecciones, la resistencia a los antibióticos amenaza la salud de todos.2 Por ello, numerosas investigaciones se dedican a comprender su expansión para controlarla mejor e incluso detenerla.
¡Los antibióticos salvan vidas! ¿Sabía que también afectan a su microbiota? ¿Sabía que un mal uso o un abuso de los antibióticos puede generar una resistencia a estos medicamentos? ¿Ha oído hablar de la Semana mundial de concienciación sobre la RAM (WAAW)? Encontrará todas las respuestas en esta página dedicada al tema:
Una resistencia a los antibióticos que pasa desapercibida
Hoy sabemos que la resistencia a los antibióticos se debe principalmente a la utilización excesiva de los antibióticos en medicina humana, pero también en la ganadería y la agricultura.2 Sin embargo, un estudio reciente nos revela un mecanismo de propagación inesperado: ¡las vacaciones y los desplazamientos profesionales a países exóticos! En efecto, los viajes internacionales favorecerían la propagación de los (sidenote:
Gen
Un gen es la unidad física y funcional básica de la herencia. Los genes están formados por ADN.
Fuente: What is a gene?MedlinePlus.gov. ) de resistencia a los antimicrobianos en el intestino. Unos investigadores reunieron a 190 viajeros holandeses, distribuidos en cuatro subgrupos según su destino. Los lugares visitados se eligieron en zonas muy afectadas por la resistencia a los antibióticos: Sudeste Asiático, Asia del Sur, África del Norte y África del Este. Estos investigadores buscaron determinar si los desplazamientos internacionales hacia estas regiones podían facilitar su diseminación hacia regiones menos afectadas. Para evaluar la presencia de estos genes en el intestino, se tomó una muestra de heces de cada participante antes y después del viaje.
Imagen
¿Una valija diplomática de la resistencia a los antibióticos en el intestino?
Gracias al empleo de una técnica de punta ( (sidenote:
Metagenómica
Método de estudio del contenido genético de muestras procedentes de entornos complejos (intestino, océano, suelos, aire, etc.) obtenidos en la naturaleza (por oposición a las muestras cultivadas en el laboratorio). Este enfoque permite describir los genes contenidos en la muestra, pero también tener una idea aproximada del potencial funcional de un entorno.
Fuente: Riesenfeld CS, Schloss PD, Handelsman J. Metagenomics: genomic analysis of microbial communities. Annu Rev Genet. 2004;38:525-52.)), el equipo constató un aumento del número de genes de resistencia a los antibióticos entre la salida y el regreso, en especial en los viajeros procedentes del Sudeste Asiático. En total, se detectaron una cincuentena de genes de resistencia a los antibióticos a raíz de los viajes. Entre ellos, genes de resistencia clásicos y bien conocidos a los antibióticos (como la familia de los β-lactámicos, las tetraciclinas, las fluoroquinolonas y otras), pero también nuevos genes nunca antes identificados.
Viajar, ¿un reto de salud pública?
Los resultados de este estudio no admiten réplica: los viajeros internacionales, colonizados por genes de resistencia durante sus viajes, podrían traer en su maleta, sin saberlo, bacterias resistentes a los antibióticos. Frente al riesgo de propagación, los autores dan la voz de alarma y resaltan la importancia de iniciar acciones con rapidez en los países especialmente afectados por la resistencia a los antibióticos. Un llamamiento que se hace eco de la campaña de la OMS.
¿Qué es la Semana mundial de concienciación sobre la RAM?
Cada año, desde 2015, la OMS organiza Semana mundial de concienciación sobre la RAM (WAAW por sus siglas en inglés), cuyo objetivo es fomentar la sensibilización sobre la resistencia global antimicrobiana.
La resistencia antimicrobiana se produce cuando las bacterias, los virus, los parásitos y los hongos cambian con el tiempo y dejan de responder a los medicamentos. Debido a la resistencia a los fármacos, los antibióticos y otros medicamentos dejan de ser efectivos y las infecciones son cada vez más difíciles o incluso imposibles de tratar, lo que aumenta el riesgo de propagación de las enfermedades, de sufrir dolencias graves y de muerte.
Esta campaña, que se desarrolla del 18 al 24 de noviembre, anima al público en general, a los profesionales de la salud y a los responsables políticos a usar los antibióticos, los antivirales, los antifúngicos y los antiparasitarios con cuidado, para prevenir la aparición de nuevas resistencias antimicrobianas.
Desde el descubrimiento de la penicilina en 1928, el uso generalizado de los antibióticos ha permitido salvar varios millones de vidas. Junto con las vacunas, los antibióticos –principales armas en la lucha contra las infecciones bacterianas– han permitido prolongar cerca de veinte años la esperanza de vida.1
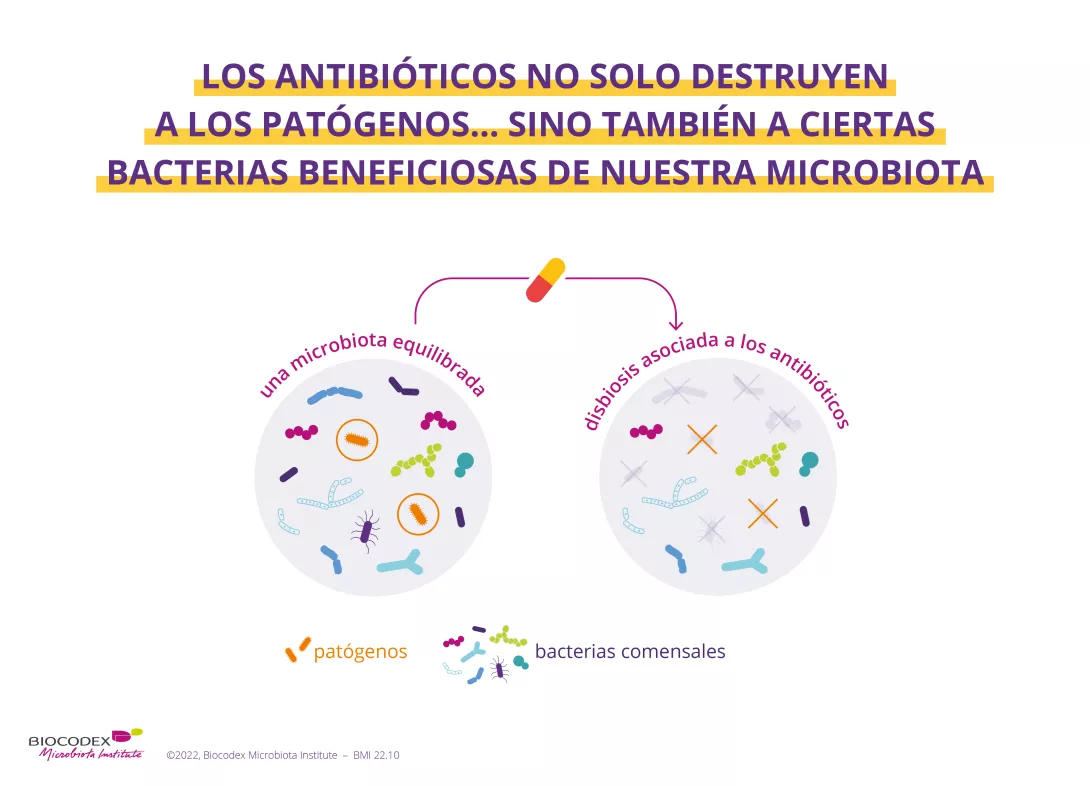
2. Los antibióticos no solo destruyen a las especies responsables de las infecciones, sino también a las bacterias beneficiosas
Intestino, vagina, pulmón, piel… Diversas partes de nuestro cuerpo contienen microorganismos (bacterias, hongos, virus). Reciben el nombre de microbiotas.2 Los antibióticos, aunque erradican los gérmenes patógenos responsables de la infección, también pueden destruir ciertas bacterias beneficiosas de nuestra microbiota y provocar un desequilibrio más o menos importante en este ecosistema. Es lo que se llama una (sidenote:
Disbiosis
La disbiosis no es un fenómeno homogéneo ya que varía en función del estado de salud de cada individuo. Se define generalmente como una alteración de la composición y funcionamiento de la microbiota, provocada por un conjunto de factores ambientales y relacionados con el individuo, que alteran el ecosistema microbiano.
Levy M, Kolodziejczyk AA, Thaiss CA, et al. Dysbiosis and the immune system. Nat Rev Immunol. 2017;17(4):219-232.)3. Todas las microbiotas del cuerpo se ven afectadas: no solo la microbiota intestinal, sino también las microbiotas cutánea4, pulmonar5, ORL6, urinaria7 y vaginal8.
3. Los antibióticos pueden tener efectos secundarios
Al producir una disbiosis, los antibióticos pueden tener consecuencias negativas para la salud. La principal complicación a corto plazo es la alteración del tránsito intestinal en algunos pacientes. Esto resulta generalmente en diarrea ya que la microbiota es menos capaz de cumplir con sus funciones protectoras. La diarrea asociada a los antibióticos suele ser de intensidad leve a moderada y su incidencia varía según la edad, el tipo de antibióticos, el contexto… Puede afectar hasta al 35%9,10,11 de los pacientes y, en los niños, el porcentaje puede llegar al 80%9. En el 10 al 20% de los casos, la diarrea se debe a una infección por Clostridioides difficile (C. difficile):11 esta bacteria coloniza la microbiota intestinal y se vuelve patógena bajo la influencia de ciertos factores (por ejemplo, la toma de antibióticos). Las consecuencias clínicas son variables y van desde una diarrea moderada hasta síntomas mucho más graves, incluso la muerte.11
35%
Puede afectar hasta al 35% de los pacientes
80%
y hasta al 80% en los niños
4. Los antibióticos pueden tener efectos crónicos cuando aparece muy temprano en la vida
La diarrea asociada a los antibióticos no es la única manifestación de la disbiosis asociada a los antibióticos. Esta última sería responsable de efectos crónicos cuando aparece muy temprano en la vida. De hecho, el periodo perinatal, que se caracteriza por el desarrollo de la microbiota intestinal y la maduración del sistema inmunitario, constituye un periodo especialmente sensible:12 la disbiosis inducida por la toma de antibióticos durante esta fase parece ser un factor de riesgo en la aparición de enfermedades crónicas (obesidad, diabetes, asma, enfermedades inflamatorias crónicas del intestino).13
5. El uso inadecuado de los antibióticos es responsable de la resistencia a los antibióticos
La resistencia a los antibióticos es el fenómeno que provoca que un tratamiento antibiótico deje de ser eficaz sobre una infección bacteriana.1 ¿A qué se debe? Los antibióticos solo son eficaces contra las bacterias y no tienen ningún efecto en los virus (por ejemplo, el de la gripe).14 Su uso inadecuado (en caso de infección vírica, por ejemplo) o excesivo, en el hombre o el animal, acelera este fenómeno. La resistencia a los antibióticos comporta hospitalizaciones más prolongadas, un mayor gasto sanitario y un aumento de la mortalidad. Por eso constituye, a escala mundial, un reto importante de salud pública.1
6. Cada año, se dedica una Semana Mundial a favorecer el uso adecuado de los antibióticos
Cada año, del 18 al 24 de noviembre, la OMS organiza la Semana mundial de concienciación sobre la RAM que pretende dar a conocer mejor el fenómeno de la resistencia a los (sidenote:
Antimicrobianos
Categoría de medicamentos que agrupa los antibióticos (activos contra bacterias), los antivirales (activos contra virus), los antiparasitarios (activos contra parásitos) y los antifúngicos (activos contra hongos)
WHO Antimicrobial Resistance; Oct 2020) y animar al público general, los profesionales sanitarios y los responsables políticos a adoptar mejores prácticas para luchar contra la aparición y la propagación de resistencias. En su calidad de experto en las microbiotas, el Biocodex Microbiota Institute se une a esta iniciativa.
Esta campaña, que tendrá lugar del 18 al 24 de noviembre, alienta al público general, a los profesionales sanitarios y a los responsables a hacer un uso razonable de los antimicrobianos para evitar el desarrollo de resistencia a los antimicrobianos.
La parte positiva de los antibióticos es que son uno de los pilares de nuestro arsenal terapéutico y salvan millones de vidas cada año. La parte negativa es que alteran nuestra microbiota y pueden tener consecuencias importantes para nuestra salud. Examinamos 6 puntos esenciales para utilizarlos con buen criterio.
Desde el descubrimiento de la penicilina en 1928, el uso generalizado de los antibióticos ha permitido salvar varios millones de vidas. Junto con las vacunas, los antibióticos –principales armas en la lucha contra las infecciones bacterianas– han permitido prolongar cerca de veinte años la esperanza de vida.1
Imagen
2. Los antibióticos no solo destruyen a las especies responsables de las infecciones, sino también a las bacterias beneficiosas
Intestino, vagina, pulmón, piel… Diversas partes de nuestro cuerpo contienen (sidenote:
Microorganismos
Organismos vivos que son demasiado pequeños para observarlos a simple vista. Incluyen bacterias, virus, hongos, arqueas, protozoos, etc. y se llaman comúnmente «microbios».
What is microbiology? Microbiology Society.) (bacterias, hongos, virus). Reciben el nombre de microbiotas.2 Los antibióticos, aunque erradican los gérmenes patógenos responsables de la infección, también pueden destruir ciertas bacterias beneficiosas de nuestra microbiota y provocar un desequilibrio más o menos importante en este ecosistema. Es lo que se llama una (sidenote:
Disbiosis
La disbiosis no es un fenómeno homogéneo ya que varía en función del estado de salud de cada individuo. Se define generalmente como una alteración de la composición y funcionamiento de la microbiota, provocada por un conjunto de factores ambientales y relacionados con el individuo, que alteran el ecosistema microbiano.
Levy M, Kolodziejczyk AA, Thaiss CA, et al. Dysbiosis and the immune system. Nat Rev Immunol. 2017;17(4):219-232.).
Imagen
Todas las microbiotas del cuerpo se ven afectadas:
3. Los antibióticos pueden tener efectos secundarios
Al producir una disbiosis, los antibióticos pueden tener consecuencias negativas para la salud. La principal complicación a corto plazo es la alteración del tránsito intestinal en algunos pacientes. Esto resulta generalmente en diarrea ya que la microbiota es menos capaz de cumplir con sus funciones protectoras. La diarrea asociada a los antibióticos suele ser de intensidad leve a moderada y su incidencia varía según la edad, el tipo de antibióticos, el contexto… Puede afectar hasta al 35%9,10,11 de los pacientes y, en los niños, el porcentaje puede llegar al 80%9. En el 10 al 20% de los casos, la diarrea se debe a una infección por Clostridioides difficile (C. difficile):11 esta bacteria coloniza la microbiota intestinal y se vuelve patógena bajo la influencia de ciertos factores (por ejemplo, la toma de antibióticos). Las consecuencias clínicas son variables y van desde una diarrea moderada hasta síntomas mucho más graves, incluso la muerte.11
35%
La diarrea asociada a antibióticos puede afectar hasta al 35% de los pacientes
80%
y hasta el 80% si los pacientes son niños
4. Los antibióticos pueden tener efectos crónicos cuando aparece muy temprano en la vida
La diarrea asociada a los antibióticos no es la única manifestación de la disbiosis asociada a los antibióticos. Esta última sería responsable de efectos crónicos cuando aparece muy temprano en la vida. De hecho, el periodo perinatal, que se caracteriza por el desarrollo de la microbiota intestinal y la maduración del sistema inmunitario, constituye un periodo especialmente sensible:12 la disbiosis inducida por la toma de antibióticos durante esta fase parece ser un factor de riesgo en la aparición de enfermedades crónicas (obesidad, diabetes, asma, enfermedades inflamatorias crónicas del intestino).13
Imagen
5. El uso inadecuado de los antibióticos es responsable de la resistencia a los antibióticos
La resistencia a los antibióticos es el fenómeno que provoca que un tratamiento antibiótico deje de ser eficaz sobre una infección bacteriana.1 ¿A qué se debe? Los antibióticos solo son eficaces contra las bacterias y no tienen ningún efecto en los virus (por ejemplo, el de la gripe).14 Su uso inadecuado (en caso de infección vírica, por ejemplo) o excesivo, en el hombre o el animal, acelera este fenómeno. La resistencia a los antibióticos comporta hospitalizaciones más prolongadas, un mayor gasto sanitario y un aumento de la mortalidad. Por eso constituye, a escala mundial, un reto importante de salud pública.1
Imagen
6. Cada año, se dedica una Semana mundial de concienciación sobre la RAM
Cada año, del 18 al 24 de noviembre, la OMS organiza la Semana mundial de concienciación sobre la RAM que pretende dar a conocer mejor el fenómeno de la resistencia a los (sidenote:
Antimicrobianos
Categoría de medicamentos que agrupa los antibióticos (activos contra bacterias), los antivirales (activos contra virus), los antiparasitarios (activos contra parásitos) y los antifúngicos (activos contra hongos)
WHO Antimicrobial Resistance; Oct 2020) y animar al público general, los profesionales sanitarios y los responsables políticos a adoptar mejores prácticas para luchar contra la aparición y la propagación de resistencias. En su calidad de experto en las microbiotas, el Biocodex Microbiota Institute se une a esta iniciativa.
Si le interesan los efectos de los antibióticos en su salud y su microbiota o si quiere saber más sobre la Semana mundial de concienciación sobre la RAM (WAAW), le recomendamos que visite esta otra página dedicada al tema:
¿Qué es la Semana mundial de concienciación sobre la RAM?
Cada año, desde 2015, la OMS organiza la Semana mundial de concienciación sobre la RAM (WAAW por sus siglas en inglés), cuyo objetivo es fomentar la sensibilización sobre la resistencia global antimicrobiana.
La resistencia antimicrobiana se produce cuando las bacterias, los virus, los parásitos y los hongos cambian con el tiempo y dejan de responder a los medicamentos. Debido a la resistencia a los fármacos, los antibióticos y otros medicamentos dejan de ser efectivos y las infecciones son cada vez más difíciles o incluso imposibles de tratar, lo que aumenta el riesgo de propagación de las enfermedades, de sufrir dolencias graves y de muerte.
Esta campaña, que se desarrolla del 18 al 24 de noviembre, anima al público en general, a los profesionales de la salud y a los responsables políticos a usar los antibióticos, los antivirales, los antifúngicos y los antiparasitarios con cuidado, para prevenir la aparición de nuevas resistencias antimicrobianas.
A la larga, podría poner en jaque un siglo de avances médicos.1 Toda una bomba sanitaria de relojería, la antibiorresistencia está en el punto de mira de la OMS, que desde 2015 organiza cada año la Semana mundial de concienciación sobre la RAM (del 18 al 24 de noviembre). El Institut du Microbiote (Instituto de la Microbiota) participa activamente en esta iniciativa con la difusión y el intercambio de contenidos exclusivos sobre el impacto de los antimicrobianos en la microbiota intestinal durante todo el mes de noviembre. Panorama general.
Por una parte, un extraordinario descubrimiento científico que permite salvar millones de vidas. Por otra parte, un uso excesivo y a veces inadecuado que puede provocar varias resistencias de los microorganismos (bacterias, virus, parásitos, hongos). La consecuencia es que los antimicrobianos, concebidos para curar, son cada vez menos eficaces y, si no se toma ninguna medida a largo plazo, corremos el riesgo de no poder curar las infecciones.
10 millones
La resistencia a los antimicrobianos sería responsable de casi 10 millones de muertes en el mundo para 2050
De esta forma, los antimicrobianos serían responsables de la muerte de casi 700 000 personas en todo el mundo cada año.2 Si nada cambia, las enfermedades infecciosas podrían convertirse, en 2050, en unas de las primeras causas de mortalidad del mundo y provocarían hasta 10 millones de muertes.2
Ante este panorama, la OMS reacciona a nivel mundial. Desde 2015, del 18 al 24 de noviembre, organiza la Semana mundial de concienciación sobre la RAM que busca concienciar sobre este fenómeno mundial y anima al gran público, a los profesionales sanitarios y a los responsables de la toma de decisiones a adoptar un uso racional de los antibióticos para evitar que la antibiorresistencia siga ganando terreno.
Toda una institución en cuanto al conocimiento sobre la microbiota, el Institut du Microbiote (Instituto de la Microbiota) participa activamente en este evento desde 2020. Durante todo el mes de noviembre, el Instituto les invita, a través de artículos, noticias y también vídeos de expertos y documentación temática, a profundizar sus conocimientos sobre el tema y a descubrir las consecuencias a medio y largo plazo de los antibióticos en la microbiota humana. ¿Un ejemplo? Pese a su eficacia reconocida contra las bacterias (y su inutilidad en caso de infección viral)3, a menudo provocan una disbiosis. Esto está relacionado con algunos problemas que todos conocemos, como la diarrea asociada a los antibióticos.
Los antibióticos, elemento clave del arsenal terapéutico moderno, han salvado millones de vidas. Sin embargo, su uso excesivo y a veces inadecuado puede conducir a la aparición de distintas formas de resistencia en los microorganismos. Cada año, la Organización Mundial de la Salud (OMS) organiza la Semana mundial de concienciación sobre la RAM (WAAW) para sensibilizar a la población sobre este problema de salud pública. Lea la página dedicada a este asunto.
¡Pero no acaba aquí! También se sospecha que los antibióticos pueden aumentar el riesgo de sufrir varias enfermedades crónicas4 (alergias, asma, obesidad, enfermedad inflamatoria crónica del intestino...), especialmente si se toman pronto durante la infancia. ¿Podemos remediarlo? ¡Sí! Solo tenemos que apostar por una prescripción pertinente para garantizar el buen uso de los antibióticos. Pero también asesorar al paciente sobre los riesgos de disbiosis asociados a un uso excesivo e inadecuado de los antibióticos. Todos responsables y movilizados para reducir la resistencia frente a los antimicrobianos.
Les presentamos al Profesor Sørensen, ganador de la bolsa internacional 2022 de la Biocodex Microbiota Foundation.
Su equipo fue el primero en lanzar un estudio de gran evergadura sobre el resistoma de 700 niños, que permitirá dar un paso agigantado en la comprensión de la evolución y diseminación de la resistencia a los antimicrobianos en el intestino humano al principio de la vida.
3 Improving Antibiotic Use. Material Developed by CDC Using CDC materials does not imply endorsement or recommendation by CDC, ATSDR, HHS or the United States Government
A la larga, podría poner en jaque un siglo de avances médicos.1 Toda una bomba sanitaria de relojería, la antibiorresistencia está en el punto de mira de la OMS, que desde 2015 organiza cada año la Semana mundial de concienciación sobre la RAM (del 18 al 24 de noviembre). El Institut du Microbiote (Instituto de la Microbiota) participa activamente en esta iniciativa con la difusión y el intercambio de contenidos exclusivos sobre el impacto de los antimicrobianos en la microbiota intestinal durante todo el mes de noviembre. Panorama general.
Por una parte, un extraordinario descubrimiento científico que permite salvar millones de vidas. Por otra parte, un uso excesivo y a veces inadecuado que puede provocar varias resistencias de los microorganismos (bacterias, virus, parásitos, hongos). La consecuencia es que los antimicrobianos, concebidos para curar, son cada vez menos eficaces y, si no se toma ninguna medida a largo plazo, corremos el riesgo de no poder curar las infecciones.
Imagen
De esta forma, los antimicrobianos serían responsables de la muerte de casi 700 000 personas en todo el mundo cada año.2 Si nada cambia, las enfermedades infecciosas podrían convertirse, en 2050, en unas de las primeras causas de mortalidad del mundo y provocarían hasta 10 millones de muertes2.
10 millones
La resistencia a los antimicrobianos seria responsable de casi 10 millones de muertes en todo el mundo para 2050
Ante este panorama, la OMS reacciona a nivel mundial. Desde 2015, del 18 al 24 de noviembre, organiza la Semana mundial de concienciación sobre la RAMque busca concienciar sobre este fenómeno mundial y anima al gran público, a los profesionales sanitarios y a los responsables de la toma de decisiones a adoptar un uso racional de los antibióticos para evitar que la antibiorresistencia siga ganando terreno.
Imagen
Toda una institución en cuanto al conocimiento sobre la microbiota, el Institut du Microbiote (Instituto de la Microbiota) participa activamente en este evento desde 2020. Durante todo el mes de noviembre, el Instituto les invita, a través de artículos, noticias y también vídeos de expertos, a descubrir las consecuencias a medio y largo plazo de los antibióticos en la microbiota humana. ¿Un ejemplo? Lo vemos con los antibióticos. Pese a su eficacia reconocida contra las bacterias (y su inutilidad en caso de infección viral)3, perturban el equilibrio de nuestra microbiota intestinal. Este desequilibrio, más conocido como (sidenote:
Disbiosis
La disbiosis no es un fenómeno homogéneo ya que varía en función del estado de salud de cada individuo. Se define generalmente como una alteración de la composición y funcionamiento de la microbiota, provocada por un conjunto de factores ambientales y relacionados con el individuo, que alteran el ecosistema microbiano.
Levy M, Kolodziejczyk AA, Thaiss CA, et al. Dysbiosis and the immune system. Nat Rev Immunol. 2017;17(4):219-232.), está relacionado con algunos problemas que todos conocemos, como la diarrea asociada a los antibióticos. ¡Pero no acaba aquí! También se sospecha que los antibióticos pueden aumentar el riesgo de sufrir varias enfermedades crónicas (alergias, asma, obesidad, enfermedad inflamatoria crónica del intestino…), especialmente si se toman pronto durante la infancia.
Imagen
¿Podemos remediarlo? ¡Sí! En primer lugar, debemos ceñirnos a un uso limitado y adecuado. Nunca debemos tomar estos fármacos sin la prescripción de un profesional sanitario. También debemos respetar la dosis, la posología, la duración del tratamiento y no compartirlos con nadie.4
¡No debemos olvidar que los antibióticos no surten efecto automáticamente!
Si le interesan los efectos de los antibióticos en su salud y su microbiota o si quiere saber más sobre la Semana mundial de concienciación sobre la RAM (WAAW), le recomendamos que visite esta otra página dedicada al tema:
¿Qué es la Semana mundial de concienciación sobre la RAM?
Cada año, desde 2015, la OMS organiza la Semana mundial de concienciación sobre la RAM (WAAW por sus siglas en inglés), cuyo objetivo es fomentar la sensibilización sobre la resistencia global antimicrobiana.
La resistencia antimicrobiana se produce cuando las bacterias, los virus, los parásitos y los hongos cambian con el tiempo y dejan de responder a los medicamentos. Debido a la resistencia a los fármacos, los antibióticos y otros medicamentos dejan de ser efectivos y las infecciones son cada vez más difíciles o incluso imposibles de tratar, lo que aumenta el riesgo de propagación de las enfermedades, de sufrir dolencias graves y de muerte.
Esta campaña, que se desarrolla del 18 al 24 de noviembre, anima al público en general, a los profesionales de la salud y a los responsables políticos a usar los antibióticos, los antivirales, los antifúngicos y los antiparasitarios con cuidado, para prevenir la aparición de nuevas resistencias antimicrobianas.
Revisión de prensa Por el Pr. Markku Voutilainen Facultad de medicina de la Universidad de Turku; gastroenterología, Hospital Universitario de Turku, Finlandia
Un estudio reciente ha demostrado que el aceite de coco puede ayudar a mantener un cuero cabelludo sano, ya que mejora su microbiota. Los investigadores compararon el efecto del aceite de coco y de un champú neutro en la microbiota bacteriana y fúngica del cuero cabelludo de 140 mujeres con y sin caspa. El cuero cabelludo de las mujeres con caspa presentaba una abundancia muy superior de la especies no caracterizadas de Malassezia, unos hongos conocidos por acelerar la aparición de la caspa y la inflamación. Por el contrario, la especie de hongo M. globosa se encontró en abundancia en el cuero cabelludo de las mujeres que no tenían caspa ni picor. El tratamiento con aceite de coco hizo que la proporción de M. globosa con respecto a otros grupos de Malassezia coincidiera con la de cueros cabelludos sanos. Aunque no se hallaron diferencias significativas entre la microbiota bacteriana del grupo sano y la del grupo con caspa, en ambos se produjo un aumento de las bacterias implicadas en el metabolismo de la biotina tras el tratamiento con aceite de coco. La biotina, una vitamina B, es fundamental para mantener la salud de la piel y del cuero cabelludo. También se sabe que la biotina reduce la inflamación. Se necesitan más estudios para comprender los mecanismos subyacentes, pero para los investigadores, el efecto positivo del aceite de coco en la composición y la función de la microbiota del cuero cabelludo es el primer paso para una recuperación duradera de un cuero cabelludo sano.
Revisión de prensa Por el Pr. Markku Voutilainen Facultad de medicina de la Universidad de Turku; gastroenterología, Hospital Universitario de Turku, Finlandia
Un estudio ha demostrado que la utilización de endolisinas recombinantes de tipo 1,4-beta-N-acetilmuramidasa codificadas en profagos de Gardnerella permite eliminar el biofilm bacteriano causante de la vaginosis bacteriana, sin dañar las bacterias beneficiosas de la microbiota vaginal. Para ello, los autores generaron varias endolisinas manipuladas genéticamente, enzimas de bacteriófagos que lisan la pared bacteriana, a través de un barajado de dominios (domain shuffling). Compararon su actividad bactericida en cepas de Gardnerella con la de endolisinas naturales. La actividad bactericida de las endolisinas recombinantes fue 10 veces mayor a la de cualquier enzima natural. Cuando se probó en un grupo de 20 cepas de Gardnerella de 4 especies diferentes (G. vaginalis, G. leopoldii, G. piotii y G. swidsinski), la endolisina más activa, denominada PM-477, demostró una eficacia superior a la de los antibióticos evaluados (metronidazol, tinidazol, clindamicina). Es más, la PM-477 no tuvo ningún efecto sobre los lactobacilos beneficiosos ni en otras especies de bacterias vaginales. Según los autores, la PM-477 es altamente selectiva para Gardnerella y mata cepas de cada una de las cuatro especies principales sin afectar a los lactobacilos beneficiosos ni a otras especies típicas de la microbiota vaginal. El efecto de la PM-477 se confirmó por microscopía en cultivos mixtos de Gardnerella y lactobacilos. Para ir más allá y analizar la eficacia de la PM-477 en un entorno fisiológico muy similar a la situación in vivo, los investigadores trataron hisopos vaginales de 15 pacientes con vaginosis bacteriana y los analizaron mediante hibridación in situ con fluorescencia (HISF). Demostraron que, en 13 de los 15 casos, la PM-477 eliminó la bacteria Gardnerella y disolvió físicamente los biofilms sin influir en la microbiota vaginal. Para los autores, las endolisinas son una prometedora alternativa terapéutica a los antibióticos para el tratamiento de la vaginosis bacteriana.